
Майская образовательная программа по генетике: Руководители
Положение о Майской образовательной программе по генетике
Образовательного центра «Сириус»
1. Общие положения
Настоящее Положение определяет порядок организации и проведения Майской образовательной программы по генетике Образовательного центра «Сириус» (далее — образовательная программа), её методическое и финансовое обеспечение.
1.1. Образовательная программа проводится в Образовательном центре «Сириус» (Образовательный Фонд «Талант и успех») с 7 по 30 мая 2021 года.
1.2. Для участия в образовательной программе приглашаются школьники 9-10 классов (на май 2021 года) из образовательных организаций всех субъектов Российской Федерации. К участию в конкурсном отборе в виде исключения могут быть допущены обучающиеся 7–8 классов, прошедшие отбор по общим правилам.
Общее количество участников образовательной программы: не более 80 человек.
1.3. К участию в образовательной программе допускаются школьники, являющиеся гражданами Российской Федерации.
1.4. Персональный состав участников образовательной программы утверждается Экспертным советом Образовательного Фонда «Талант и успех» по направлению «Наука».
1.5. Научно-методическое и кадровое сопровождение профильной образовательной программы осуществляют Биологический факультет МГУ имени М.В. Ломоносова, Центр педагогического мастерства г. Москвы, сотрудники вузов и институтов РАН и ФМБА, имеющие опыт работы в области классической или молекулярной генетики и необходимую педагогическую квалификацию.
1.6. В связи с целостностью и содержательной логикой образовательной программы, интенсивным режимом занятий и объемом академической нагрузки, рассчитанной на весь период пребывания обучающихся в Образовательном центре «Сириус», не допускается участие школьников в отдельных мероприятиях или части образовательной программы: исключены заезды и выезды школьников вне сроков, установленных Экспертным советом Фонда по направлению «Наука».
1.7. В случае нарушений правил пребывания в Образовательном центре «Сириус» или требований настоящего Положения решением Координационного совета участник образовательной программы может быть отчислен с образовательной программы.
1.8. В целях создания более широких возможностей посещения Образовательного центра «Сириус» допускается участие школьников в течение учебного года (с июля 2020 г. по июнь 2021 г.) не более чем в двух образовательных программах по направлению «Наука» (по любым профилям, включая проектные образовательные программы).
2. Цели и задачи образовательной программы
2.1. Образовательная программа ориентирована на выявление одаренных школьников, интересующихся генетикой, развитие их творческих способностей, знакомство с практическим и исследовательским применением современных генетических технологий, а также повышение образовательного уровня школьников в области генетики и смежных дисциплин.
2.2. Задачи образовательной программы:
– знакомство школьников с актуальным положением в областях молекулярной и медицинской генетики;
– развитие практических способностей учащихся в области современных генетических методов;
– развитие умений и навыков решения задач по генетике и молекулярной генетике;
– развитие у школьников навыков критического мышления и формирование у них представлений о возможностях современных генетических технологий;
– популяризация генетики как науки.
3. Порядок отбора участников образовательной программы
3.1. Отбор участников осуществляется Координационным советом, формируемым Руководителем Образовательного Фонда «Талант и успех», на основании требований, изложенных в настоящем Положении, а также в Порядке отбора школьников на профильные образовательные программы Фонда по направлению «Наука».
3. 2. Для участия в конкурсном отборе необходимо пройти регистрацию на официальном сайте Образовательного центра «Сириус».
2. Для участия в конкурсном отборе необходимо пройти регистрацию на официальном сайте Образовательного центра «Сириус».
Регистрация будет открыта до 5 февраля 2021 года.
Не зарегистрировавшиеся школьники к участию в образовательной программе не допускаются.
3.4. Отбор участников осуществляется в два этапа. Первый этап — дистанционный учебно-отборочный курс в системе Сириус.Онлайн. Второй этап — заключительный очный отборочный тур (проводится в регионах).
3.4.1. С 25 января по 6 марта 2021 года для зарегистрировавшихся школьников будет организован дистанционный учебно-отборочный курс. Информация о курсе размещается в личном кабинете участника после его регистрации.
3.4.3. В рамках дистанционного учебно-отборочного курса оценивается успешность освоения учебного материала, а также результат, показанный на дистанционном тестировании, проводящемся в рамках курса. Дистанционное тестирование состоится 6 марта 2021 года.
Дистанционное тестирование состоится 6 марта 2021 года.
3.4.4. По итогам обучения в дистанционном учебно-отборочном курсе и финального дистанционного тестирования будут определены участники заключительного очного отборочного тура, который пройдет на площадках в субъектах Российской Федерации
3.4.5. Список школьников, допущенных к участию в заключительном очном отборочном туре, будет опубликован на сайте Образовательного центра «Сириус» не позднее 10 марта 2021 года.
3.4.6. Регламент проведения заключительного очного отборочного тура, места и время проведения этого тура в регионах будут опубликованы на сайте Образовательного центра «Сириус» не позднее 12 марта 2021 года.
3.5. Олимпиады, конкурсы и турниры любого уровня не дают права внеконкурсного зачисления на программу. Победители и призеры Московской олимпиады школьников по генетике 2019/2020 учебного года получают при регистрации на программу бонусный балл на дистанционном этапе, не превышающий 20% для победителей и 10% для призеров от максимальной суммы баллов за отбор дистанционного этапа.
3.6. В образовательной программе могут принять участие не более 30 школьников от одного субъекта Российской Федерации. В случае прохождения более 30 школьников от одного региона через первичный проходной балл для данного региона Координационный совет программы принимает вторичный проходной балл, допускающий прохождение не более 30 школьников.
3.7. Учащиеся, отказавшиеся от участия в образовательной программе, могут быть заменены на следующих за ними по рейтингу школьников (по итогам заключительного очного отборочного тура). Решение о замене участников принимается Координационным советом программы. Внесение изменений в список участников программы происходит до 27 апреля 2021 года.
3.8. Список участников образовательной программы будет опубликован на сайте Образовательного центра «Сириус» не позднее 1 апреля 2021 года.
4. Аннотация образовательной программы
Аннотация образовательной программы
Образовательная программа включает в себя лекционные, семинарские и практические занятия по классической, популяционной, молекулярной и медицинской генетике, лекции ведущих российских специалистов в области генетики, тематические экскурсии, общеобразовательные, культурно-досуговые и спортивно-оздоровительные мероприятия.
5. Финансирование образовательной программы
Оплата проезда, пребывания и питания школьников — участников образовательной программы осуществляется за счет средств Образовательного Фонда «Талант и успех».
ДНК – молекулярная основа наследственности. Каждая молекула ДНК является двойной цепочкой из нуклеотидов (полинуклеотидов, соединяющихся посредине, образуя двойную спираль).  Генетика (с стард.грец. genetikos – порождать и genesis-рождение) – область биологии; наука о генах, наследственности и вариативности организмов. Тот факт, что живые существа наследуют черты своих родителей, был использован еще в доисторические времена для улучшения производительности злаковых культур и животных путем селективного разведения. Однако, современная генетика, которая стремиться понять процесс наследования, реально началась только с работы Грегора Менделя в середине девятнадцатого века. Несмотря на то, что он не знал физических основ наследственности, Мендель заметил, что организмы наследуют черты через особые дискретные единицы наследственности, которые сегодня мы называем генами. Гены являются участками в ДНК – молекула, состоящая из цепи четырех различных типов нуклеотидов – последовательность которых и есть генетической информацией, которую наследуют организмы. В подавляющем большинстве случаев ДНК присутствуют в форме двойных основ, с нуклеотидами на каждой основе, комплементарно дополняющих друг друга. Последовательности нуклеотидов в генах транслируются клетками для выработки цепи аминокислот и синтеза белков – порядок аминокислот в белке соответствует порядку нуклеотидов в гене. Эта связь между последовательностью нуклеотидов и последовательность аминокислот называется генетическим кодом. Аминокислоты в белке определяют, каким образом они составляют трехмерную форму, которая, в свою очередь, отвечает за функцию белка. Белки выполняют практически все функции необходимые для жизнедеятельности клеток. При изменение последовательности оснований в том участке молекулы ДНК, которая несет генетическую информацию (гене) может изменить последовательность аминокислот белка, изменив его форму и функции, что часто приводит к негативному влиянию на функционирование клеток и организма в целом. Причем в большинстве случаев происходит изменение только одного нуклеотида. 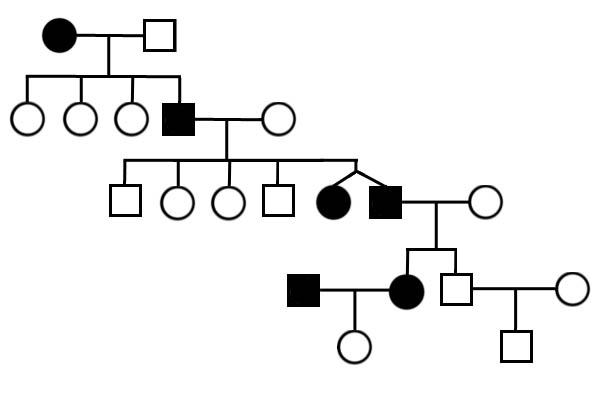 Часто это вызывает какое-то определенное генетическое заболевание, которое может передаваться по наследству. Часто это вызывает какое-то определенное генетическое заболевание, которое может передаваться по наследству.
Хотя генетические особенности играют чрезвычайно большую роль в появлении, развитии, функционировании и поведении организмов, конечный результат (совокупность признаков и особых свойств организма) является сочетанием генетических факторов с условиями в которых происходит развитие организма. Например, наследственность играет значительную роль в формировании такого признака, как рост человека, однако, особенности питания и другие условия (например физическая активность, специфические упражнения и т.д.), могут в значительной степени модулировать указанный признак в зависимости от условий.
Работы Грегора Менделя и классическая генетика
Важность работы Менделя не получила широкого понимания аж до 1890-х, когда после его смерти другие ученые, работая над подобными проблемами, вновь обратили внимание на его исследования. Уильям Бэтсон, сторонник работы Менделя, в 1905 году предложил название новой научной дисциплины – Генетика.
После возвращения к результатам исследований Менделя, ученые пытались определить, какие именно молекулы в клетке отвечают за наследственность. В 1910 году Томас Хант Морган утверждал, что гены находятся на хромосомах, опираясь на наблюдения сцепленного наследования у дрозофилы. В 1913 его ученик Альфред Стертевант использовал феномен генетической связи, чтобы показать, что гены расположены линейно на хромосоме.
Особенности наследования На фундаментальном уровне наследования в организмах происходит с помощью определенных дискретных признаков, которые однозначно определяются генами.  Это свойство впервые была обнаружена Грегором Менделем, который изучал сегрегации наследственных признаков на примере гороха. В своих экспериментах, где он изучал особенности цвета цветка, Мендель заметил, что цветы каждого гороха были или фиолетовые или белые – и никогда не наблюдалось наличие промежуточного цвета. Это отличие, наличие различных вариантов одного и того же гена называются аллелями. Это свойство впервые была обнаружена Грегором Менделем, который изучал сегрегации наследственных признаков на примере гороха. В своих экспериментах, где он изучал особенности цвета цветка, Мендель заметил, что цветы каждого гороха были или фиолетовые или белые – и никогда не наблюдалось наличие промежуточного цвета. Это отличие, наличие различных вариантов одного и того же гена называются аллелями.В случае с горохом, который относится к диплоидному виду, каждое растение имеет две аллели данного гена, где одна аллель передается от каждого из родителей. Многие виды, включая человека, имеют именно эту схему наследования. Диплоидные организмы с двумя копиями одной и той же аллели определенного гена, называются гомозиготными, а организмы с двумя разными аллелями определенного гена называются гетерозиготными. Набор аллелей для данного организма, называется его генотипом, а наблюдаемая характеристика или признак организма называют его фенотип.  Когда говорят, что данный организм гетерозиготный по гену, часто одну аллель указывают как доминирующую (доминантную), поскольку ее качества преобладают в фенотипе организма, в то время как другие аллели называются рецессивными, поскольку их качества могут отсутствовать и не наблюдаться. Некоторые аллели не имеют полного доминирования, а взамен имеют неполное доминирование промежуточного фенотипа, или т.н. кодоминантность – обе черты являются доминантными одновременно, и обе черты присутствуют в фенотипе. Когда говорят, что данный организм гетерозиготный по гену, часто одну аллель указывают как доминирующую (доминантную), поскольку ее качества преобладают в фенотипе организма, в то время как другие аллели называются рецессивными, поскольку их качества могут отсутствовать и не наблюдаться. Некоторые аллели не имеют полного доминирования, а взамен имеют неполное доминирование промежуточного фенотипа, или т.н. кодоминантность – обе черты являются доминантными одновременно, и обе черты присутствуют в фенотипе.Когда пара организмов размножается половым путем, их потомки случайно наследуют один из двух аллелей от каждого из родителей. Наблюдение дискретного наследования и сегрегация аллелей в общем известны как первый закон Менделя, или закон сегрегации (закон единообразия гибридов первого поколения).
Взаимодействие нескольких генов
Организмы имеют тысячи генов, а во время полового размножения ассортимент этих генов в основном является независимым, то есть их наследования происходит случайным образом без связи между ними. Это означает, что наследование аллелей для желтого или зеленого цвета горошка не имеет отношения к наследованию аллелей для белого или фиолетового цвета цветов. Этот феномен, известный как “Второй закон Менделя”, или “Закон независимого наследования” (закон расщепления признаков), означает, что аллели разных генов перемешиваются между родителями для формирования потомков с различными комбинациями.  Некоторые гены не могут быть унаследованы отдельно, поскольку для них предназначена определенная генетическая связь, которая обсуждается в дальнейшем в статье. Некоторые гены не могут быть унаследованы отдельно, поскольку для них предназначена определенная генетическая связь, которая обсуждается в дальнейшем в статье.Часто разные гены могут взаимодействовать таким образом, что они влияют на одну и ту же характерную черту. Например, в пупочнике весеннем (Omphalodes verna) существует ген из аллелей, определяющих цвет цветка: голубой или пурпурный. Однако другой ген контролирует или вообще имеет цветок цвет либо он белый. Когда растение имеет две копии белой аллели, его цветы являются белыми, независимо от того первый ген имел голубую или пурпурную аллель. Это взаимодействие между генами, называется эпистаз – активность одного гена находится под влиянием вариаций других генов. Многие признаки не являются дискретными чертами (например, фиолетовые или белые цветки), но зато есть непрерывными чертами (например, человеческий рост и цвет кожи). Этот комплекс признаков является следствием наличия многих генов. Влияние этих генов является связующим звеном различных степеней влияния окружающей среды на организмы.  Наследственность – это степень вклада генов организма к комплексу характерных черт. Измерение наследственности черт является относительным – в среде которая часто изменяется, она имеет большее влияние на общую смену характерных признаков. Например, в Соединенных Штатах рост человека является комплексной чертой с вероятностью наследования 89%. Однако, в Нигерии, где люди имеют существенную разницу в возможностях доступа к хорошему питания и здравоохранению, вероятность наследования такого признака как рост всего 62%. Наследственность – это степень вклада генов организма к комплексу характерных черт. Измерение наследственности черт является относительным – в среде которая часто изменяется, она имеет большее влияние на общую смену характерных признаков. Например, в Соединенных Штатах рост человека является комплексной чертой с вероятностью наследования 89%. Однако, в Нигерии, где люди имеют существенную разницу в возможностях доступа к хорошему питания и здравоохранению, вероятность наследования такого признака как рост всего 62%.
Воспроизведение Когда происходит деление клеток, их геном полностью копируется, и каждая дочерняя клетка наследует один полный набор генов. Этот процесс называется митозом – простейшая форма воспроизведения и основа для вегетативного (бесполого) размножения. Вегетативное размножение может также происходить и в многоклеточных организмах, создавая потомков, которые наследуют геном от одного отца. Отпрысков, которые являются генетически идентичными с их родителями, называют клонами.  Эукариотные организмы часто используют половое размножение для получения потомства, имеющие смешанный генетический материал, унаследованный от двух разных отцов. Процесс полового размножения меняется (чередуется) в зависимости от типа, который содержит одну копию генома (гаплоидный) и двойную копию (диплоидный). Гаплоидные клетки образуются в результате мейоза и сливаясь с другой гаплоидной клеткой генетический материал для создания диплоидной клетки с парными хромосомами (напр. слияние яйцеклетки (гаплоидная клетка) и сперматозоида (гаплоидная клетка)) вызывает образование зиготы. Диплоидные клетки путем деления образуют гаплоидные клетки, без воспроизведения их ДНК, для создания дочерних клеток, которые случайно наследуют одну из каждой пары хромосом. Большинство животных и многие растения являются диплоидными организмами на протяжении большей части своей жизни, с гаплоидной формой, которая характерна только для одной клетки – гаметы. Несмотря на то, что они не используют гаплоидный / диплоидный способ полового размножения бактерии имеют много способов получения новой генетической информации (то есть для изменчивости).  Некоторые бактерии могут пройти конъюгацию, передавая небольшой круговой фрагмент ДНК другой бактерии. Бактерии могут также принимать чужеродные фрагменты ДНК из окружающей среды и интегрировать их в свой геном, этот феномен, известный как трансформация. Этот процесс называют также горизонтальным переносом генов – передача фрагментов генетической информации между организмами, которые не связанны между собой. Некоторые бактерии могут пройти конъюгацию, передавая небольшой круговой фрагмент ДНК другой бактерии. Бактерии могут также принимать чужеродные фрагменты ДНК из окружающей среды и интегрировать их в свой геном, этот феномен, известный как трансформация. Этот процесс называют также горизонтальным переносом генов – передача фрагментов генетической информации между организмами, которые не связанны между собой.
|
 Каждая основа выступает в качестве шаблона для создания новой основы-партнера – это физический метод копирования генов, которые могут быть унаследованы.
Каждая основа выступает в качестве шаблона для создания новой основы-партнера – это физический метод копирования генов, которые могут быть унаследованы. В своей работе “Versuche über Pflanzenhybriden” (“Опыты над гибридизацией растений”), опубликованной в 1865 году Naturforschender Verein (Общество по исследованию природы), в городе Брно (современная Чехия, тогда территория Австрийской Империи), Мендель проследил порядок наследования определенных признаков для гороха и правильно описал их математически. Хотя описанный тип наследования может наблюдаться только для нескольких признаков, в работах Менделя выдвигается мнение, что наследственность является дискретной и постоянной, а не приобретенной и, что характер наследования многих признаков может быть объяснен и описан с помощью простых математических правил и пропорций.
В своей работе “Versuche über Pflanzenhybriden” (“Опыты над гибридизацией растений”), опубликованной в 1865 году Naturforschender Verein (Общество по исследованию природы), в городе Брно (современная Чехия, тогда территория Австрийской Империи), Мендель проследил порядок наследования определенных признаков для гороха и правильно описал их математически. Хотя описанный тип наследования может наблюдаться только для нескольких признаков, в работах Менделя выдвигается мнение, что наследственность является дискретной и постоянной, а не приобретенной и, что характер наследования многих признаков может быть объяснен и описан с помощью простых математических правил и пропорций. (Прилагательное генетический, происходит от греческого слова genesis-«γένεσις, происхождения, а то и от слова genno-γεννώ,” дать жизнь, родить “, предшествует существительному, и впервые был использован в биологии в 1860 году). Бэтсон популяризировал использование слова генетика для описания науки наследования в своей вступительной речи на Третьей международной конференции по гибридизации растений в Лондоне (Англия) в 1906 году.
(Прилагательное генетический, происходит от греческого слова genesis-«γένεσις, происхождения, а то и от слова genno-γεννώ,” дать жизнь, родить “, предшествует существительному, и впервые был использован в биологии в 1860 году). Бэтсон популяризировал использование слова генетика для описания науки наследования в своей вступительной речи на Третьей международной конференции по гибридизации растений в Лондоне (Англия) в 1906 году. Результаты исследования, полученные Фрэнсисом Гальтон в 1889 году, показывают взаимосвязь между ростом потомков и средним ростом их родителей. Однако корреляция не является абсолютной и присутствуют значительные отклонения от генетической изменчивости в росте потомков, что свидетельствует о том, что окружающая среда является также важным фактором этого признака.
Результаты исследования, полученные Фрэнсисом Гальтон в 1889 году, показывают взаимосвязь между ростом потомков и средним ростом их родителей. Однако корреляция не является абсолютной и присутствуют значительные отклонения от генетической изменчивости в росте потомков, что свидетельствует о том, что окружающая среда является также важным фактором этого признака.МОЛЕКУЛЯРНАЯ ГЕНЕТИКА
Достижения в области генетики и молекулярной биологии в последние годы позволили значительно прояснить понимание патогенеза гематологических заболеваний. Не подлежит сомнению, что в основе всех опухолевых заболеваний крови лежат нарушения нормального функционирования генетической структуры клетки.
По рекомендации Всемирной Организации Здравоохранения (2000 г.), детекция генетических аномалий является обязательным условием для установления диагноза целого ряда гемобластозов. Более того, выявление определенных транслокаций и мутаций позволяет предполагать особенности течения заболевания, судить о прогнозе и подбирать адекватную терапию.
Более того, выявление определенных транслокаций и мутаций позволяет предполагать особенности течения заболевания, судить о прогнозе и подбирать адекватную терапию.
Особое значение для лечения приобрело развитие так называемой «прицельной терапии», позволяющее проводить коррекцию нарушений на генетическом или биохимическом уровне. Самыми яркими примерами лекарственных препаратов, применяемых как «прицельная терапия», являются полностью трансретиноевая кислота (АТРА) и иматиниба мезилат (Гливек), позволившие внести революционные изменения в терапию острого промиелоцитарного лейкоза и хронического миелолейкоза.
В связи с этим, понимание основ молекулярной генетики, методов молекулярной диагностики, а также цитогенетической терминологии стало необходимым для работы практических гематологов.
Статьи по молекулярной генетике:
Прогностические и предсказывающие маркеры для новых видов иммунотерапии
Mahoney KM, Atkins MB
МiR-SNP в качестве маркеров токсичности и клинических исходов при лимфоме Ходжкина
Navarro A, Muñoz C, Gaya A, Díaz-Beyá M, Gel B, Tejero R, Díaz T, Martinez A, Monzó M
Печеночные порфирии в России: мутационный анализ
Селиванова Д. С., Сурин В.Л., Лучинина Ю.А., Финк О.С., Пустовойт Я.С., Карпова И.В., Кравченко С.К., Пивник А.В.
С., Сурин В.Л., Лучинина Ю.А., Финк О.С., Пустовойт Я.С., Карпова И.В., Кравченко С.К., Пивник А.В.
Хронический лимфолейкоз трансформируется в синдром Рихтера двумя основными генетическими путями
Chigrinova E, Rinaldi A, Kwee I
Интегративный полный анализ генома выявил экспрессию черт, предсказывающих трансформацию фолликулярной лимфомы
Brodtkorb M, Lingjærde OC, Huse K, Trøen G, Hystad M, Hilden VI, Myklebust JH, Leich E, Rosenwald A, Delabie J, Holte H, Smeland EB
Физика будущего (глава из книги)
М. Каку
Смешанный фенотип острого лейкоза (MPAL): результаты лечения 100 пациентов, клинические и лабораторные особенности по классификации WHO 2008
Matutes E, Pickl WF, Van’t Veer M, Morilla R, Swansbury J, Strobl H, Attarbaschi A, Hopfinger G, Ashley S, Bene MC, Porwit A, Orfao A, Lemez P, Schabath R, Ludwig WD
Лимфопролиферативные синдромы, связанные с Х хромосомой: братья или дальние родственники?
Filipovich AH, Zhang K, Snow AL, Marsh RA
13q14 делеция при ХЛЛ вовлекает сотрудничающие опухолевые супрессоры
Palamarchuk A, Efanov A, Nazaryan N, Santanam U, Alder H, Rassenti L, Kipps T, Croce CM, Pekarsky Y
Гиперхолестеринемия стимулирует мобилизацию клеток костного мозга, воздействуя на ось SDF-1:CXCR4
Gomes AL, Carvalho T, Serpa J, Torre C, Dias S
IRF4 полиморфизм rs872071 и риск лимфомы Ходжкина
Broderick P, Cunningham D, Vijayakrishnan J, Cooke R, Ashworth A, Swerdlow A, Houlston R
Иммуномодулирующий эффект 5-азацитидина (5-azaC): потенциальная роль в трансплантации
Sanchez-Abarca LI, Gutierrez-Cosio S, Santamaria C, Caballero-Velazquez T, Blanco B, Herrero-Sanchez C, Garcia JL, Carrancio S, Hernandez-Campo P, Gonzalez FJ, Flores T, Ciudad L, Ballestar E, Del Canizo C, San Miguel JF, Perez-Simon JA
Генетические факторы 18 хромосомы мышей ответственны за герминогенные тестикулярные опухоли (ГТО) и возможность деривации в эмбриональные стволовые клетки
Anderson PD, Nelson VR, Tesar PJ, Nadeau JH
Регуляция JAK2 посредством miR-135a: прогностическое значение для классической ходжкинской лимфомы
Navarro A, Diaz T, Martinez A, Gaya A, Pons A, Gel B, Codony C, Ferrer G, Martinez C, Montserrat E, Monzo M
Фармакогенетическое исследование лимфомы Ходжкина показывает воздействие полиморфизма UGT1A1 на прогноз для пациента
Ribrag V, Koscielny S, Casasnovas O, Cazeneuve C, Brice P, Morschhauser F, Gabarre J, Stamatoullas A, Lenoir G, Salles G
c-Myc–контролируемая клеточная дифференциация мегакариоцит-эритроцитарных предшественников
Guo Y, Niu C, Breslin P, Tang M, Zhang S, Wei W, Kini AR, Paner GP, Alkan S, Morris SW, Diaz M, Stiff PJ, Zhang J
Двойная терапевтическая эффективность винбластина как уникального химиотерапевтического агента, вероятно, стимулирует созревание дендроцитов
Tanaka H, Matsushima H, Nishibu A, Clausen BE, Takashima A
Мутация гена тромбомодулина при атипичном гемолитико-уремическом синдроме
Delvaeye M, Noris M, De Vriese A, Esmon CT, Esmon NL, Ferrell G, Del-Favero J, Plaisance S, Claes B, Lambrechts D, Zoja C, Remuzzi G, Conway EM
Анализ микромножества лимфатической ткани показывает определенную для стадии генную экспрессию при ВИЧ 1 инфекции
Li Q, Smith AJ, Schacker TW, Carlis JV, Duan L, Reilly CS, Haase AT
Изучение кислород-направленных путей в физиологии и патологии эритроцитоза
Semenza GL
Общие генетические варианты в генах кандидатах и риск семейной лимфоидной малигнизации
Liang XS, Caporaso N, McMaster ML, Ng D, Landgren O, Yeager M, Chanock S, Goldin LR
Терапевтическое планирование при MLL
Liedtke M, Cleary ML
Мутация MPLW515L при остром мегакариобластном лейкозе
Hussein K, Bock O, Theophile K, Schulz-Bischof K, Porwit A, Schlue J, Jonigk D, Kreipe H
Предсказатели первичной резистентности к иматинибу при хроническом миелоидном лейкозе отличны от факторов при вторичной резистентности
Zhang WW, Cortes JE, Yao H, Zhang L, Reddy NG, Jabbour E, Kantarjian HM, Jones D
Рациональное объединенное планирование фосфодиэстеразы 4B и SYK при диффузной В-крупноклеточной лимфоме
Kim SW, Rai D, McKeller MR, Aguiar RC
Специфический кариотип микроРНК при хроническом лимфоцитарном лейкозе
Visone R, Rassenti LZ, Veronese A, Taccioli C, Costinean S, Aguda BD, Volinia S, Ferracin M, Palatini J, Balatti V, Alder H, Negrini M, Kipps TJ, Croce CM
Обширное внутриклоновое разнообразие в подгруппе со стереотипными рецепторами IGHV4-34 у пациентов с хроническим лимфоцитарным лейкозом: значения для продолжающихся взаимодействий с антигеном
Sutton LA, Kostareli E, Hadzidimitriou A, Darzentas N, Tsaftaris A, Anagnostopoulos A, Rosenquist R, Stamatopoulos K
Гематлас: характеристика экспрессии генов в дифференцированных клетках крови человека
Watkins NA, Gusnanto A, de Bono B, De S, Miranda-Saavedra D, Hardie DL, Angenent WG, Attwood AP, Ellis PD, Erber W, Foad NS, Garner SF, Isacke CM, Jolley J, Koch K, Macaulay IC, Morley SL, Rendon A, Rice KM, Taylor N, Thijssen-Timmer DC, Tijssen MR, van der Schoot CE, Wernisch L, Winzer T, Dudbridge F, Buckley CD, Langford CF, Teichmann S, Göttgens B, Ouwehand WH
Индуцированные плюрипотентные стволовые клетки открывают новый подход к терапии талассемии и серповидно-клеточной анемии
Ye L, Chang JC, Lin C, Sun X, Yu J, Kan YW
Ассоциация амплификации MLL с плохим результатом при остром миелоидном лейкозе
Maitta RW, Cannizzaro LA, Ramesh KH
Метилтрансфераза при миелодиспластическом синдроме: виновна или невиновна?
Bruno Quesnel
Контроль минимальной резидуальной болезни при остром миелобластном лейкозе с мутациями NPM1
Papadaki C, Dufour A, Seibl M, Schneider S, Bohlander SK, Zellmeier E, Mellert G, Hiddemann W, Spiekermann K
Благотворное влияние мутаций CEBPA у пациентов с острым миелобластным лейкозом (AML) наблюдается только в отсутствии цитогенетических отклонений и внутренней дупликации FLT3 (FLT3-ITD)
Renneville A, Boissel N, Gachard N, Naguib D, Bastard C, de Botton S, Nibourel O, Pautas C, Reman O, Thomas X, Gardin C, Terre C, Castaigne S, Preudhomme C, Dombret H
Мониторинг и интерпретация результатов молекулярного ответа при ХМЛ
A. Г. Туркина
Г. Туркина
Мутация JAK2V617F при ранней диагностике Ph-миелопролиферативных заболеваний у пациентов с венозными тромбоэмболиями: результаты метаанализа
Dentali F, Squizzato A, Brivio L, Appio L, Campiotti L, Crowther M, Grandi AM, Ageno W
B-клеточные клоны как ранние маркеры для хронического лимфоцитарного лейкоза
Landgren O, Albitar M, Ma W, Abbas Fi, Hayes RB, Ghia P, Marti GE, Caporaso NE
Три новых пути хромосомной реаранжировки, вовлекающие MLL, замаскированный t(9;19) (p11;p13) у младенца с острым миелоидным лейкозом
de Jesus Marques-Salles T, Liehr T, Mkrtchyan H, Raimondi SC, Tavares de Souza M, de Figueiredo AF, Rouxinol S, Jordy Macedo FC, Abdelhay E, Santos N, Macedo Silva ML
Прогностическое воздействие мутаций WT1 при цитогенетически нормальном остром миелоидном лейкозе (AML): исследование немецко-австрийской группы по изучению AML (AMLSG)
Gaidzik VI, Schlenk RF, Moschny S, Becker A, Bullinger L, Corbacioglu A, Krauter J, Schlegelberger B, Ganser A, Dohner H, Dohner K
В естественных условиях пониженная регуляция микроРНК-29с (microRNA-29c) и микроРНК-223 (microRNA-223) имеют значение в развитии хронического лимфоцитарного лейкоза и улучшают стратификацию риска болезни
Stamatopoulos B, Meuleman N, Haibe-Kains B, Saussoy P, Van den Neste E, Michaux L, Heimann P, Martiat P, Bron D, Lagneaux L
Эпигенетическое регулирование микроРНК (MicroRNAs) при остром лимфобластном лейкозе
Roman-Gomez J, Agirre X, Jiménez-Velasco A, Arqueros V, Vilas-Zornoza A, Rodriguez-Otero P, Martin-Subero I, Garate L, Cordeu L, José-Eneriz ES, Martin V, Castillejo JA, Bandrés E, Calasanz MJ, Siebert R, Heiniger A, Torres A, Prosper F
IL-17 способствует CD4-установленной болезни «трансплантат против хозяина»
Kappel LW, Goldberg GL, King CG, Suh DY, Smith OM, Ligh C, Holland AM, Grubin J, Mark NM, Liu C, Iwakura Y, Heller G, van den Brink MR
Новый молекулярный механизм первичного сопротивления ингибиторам FLT3-киназы при острой миелоидной лейкемии
Breitenbuecher F, Markova B, Kasper S, Carius B, Stauder T, Bohmer FD, Masson K, Ronnstrand L, Huber C, Kindler T, Fischer T
Молекулярное распознавание острого миелоидного лейкоза при использовании аптамеров
Sefah K, Tang ZW, Shangguan DH, Chen H, Lopez-Colon D, Li Y, Parekh P, Martin J, Meng L, Phillips JA, Kim YM, Tan WH
Ингибиторы гистон диацетилазы: эффекты апоптоза и клинические значения (обзор)
Emanuele S, Lauricella M, Tesoriere G
Мутация NOTCh2/FBXW7 идентифицирует большую подгруппу с благоприятным результатом при Т-клеточном остром лимфобластном лейкозе взрослых: исследование GRAALL
Asnafi V, Buzyn A, Le Noir S, Baleydier F, Simon A, Beldjord K, Reman O, Witz F, Fagot T, Tavernier E, Turlure P, Leguay T, Huguet F, Vernant JP, Daniel F, Bene MC, Ifrah N, Thomas X, Dombret H, Macintyre E
Увеличенние риска развития полицитемии, тромбоцитемии и миелофиброза среди родственников первой степени пациентов с миелопролиферативными заболеваниями в Швеции
Landgren O, Goldin LR, Kristinsson SY, Helgadottir EA, Samuelsson J, Björkholm M
Критическая роль DAP10 и DAP12 в CD8+ Т-клеточном лейкозе из больших гранулярных лимфоцитов
Chen X, Bai F, Sokol L, Zhou J, Ren A, Painter JS, Liu J, Sallman DA, Chen YA, Yoder JA, Djeu JY, Loughran TP Jr, Epling-Burnette PK, Wei S
Эпигенетические нарушения при гемобластозах
Генетические основы гемобластозов
ЭКСПЕРИМЕНТАЛЬНЫЕ СТАТЬИ |
|
Дедков В. С. Определение специфичности ДНК-метилтрансферазы M. Bsc4I в клеточном лизате при помощи блокирования эндонуклеаз рестрикции и компьютерного моделирования | Контакты, аннотация, ключевые слова, литература С. Определение специфичности ДНК-метилтрансферазы M. Bsc4I в клеточном лизате при помощи блокирования эндонуклеаз рестрикции и компьютерного моделирования | Контакты, аннотация, ключевые слова, литература |
3 |
| Иванов М. К., Брагин А. Г., Прасолова М. А., Ведерников В. Е., Дымшиц Г. М. Разработка неконкурентного экзогенного внутреннего контроля ПЦР с детекцией в режиме реального времени по принципу UFA | Контакты, аннотация, ключевые слова, литература | 8 |
| Балахонов С. В., Шестопалов М. Ю., Романова И. Ф. Результаты VNTR-анализа по локусу (5′-CAAA-3′)n штаммов Yersinia pestis из активных природных очагов чумы Сибири |Контакты, аннотация, ключевые слова, литература | 14 |
Савельева Н. В., Загрядская Ю. Е., Климашевская С. В., Пузырев В. Ф., Бурков А. Н., Обрядина А. П., Уланова Т. И. Сканирование диагностически значимых антигенных областей основного белка Chlamydia trachomatis MOMP с помощью серии перекрывающихся рекомбинантных белков | Контакты, аннотация, ключевые слова, литература В., Загрядская Ю. Е., Климашевская С. В., Пузырев В. Ф., Бурков А. Н., Обрядина А. П., Уланова Т. И. Сканирование диагностически значимых антигенных областей основного белка Chlamydia trachomatis MOMP с помощью серии перекрывающихся рекомбинантных белков | Контакты, аннотация, ключевые слова, литература |
17 |
| Ерошенко Г. А., Видяева Н. А., Одиноков Г. Н., Куклева Л. М., Краснов Я. М., Гусева Н. П., Кутырев В. В. Структурно-функциональный анализ гена araC у штаммов Yersinia pestis различного происхождения | Контакты, аннотация, ключевые слова, литература | 21 |
| Костина Л. В., Непоклонов Е. А., Забережный А. Д., Южаков А. Г., Козлов А. Ю. Картирование антигенных детерминант белка E2 вируса классической чумы свиней с помощью синтетических пептидов | Контакты, аннотация, ключевые слова, литература | 26 |
Шарапова Н. Е., Котнова А. П., Галушкина З. М., Полетаева Н. Н., Лаврова Н. В., Аксенова Е. И., Семихин А. С., Карягина А. С., Лунин В. Г. Получение и характеристика коллагенсвязывающих доменов из фактора фон Виллебранда человека | Контакты, аннотация, ключевые слова, литература Е., Котнова А. П., Галушкина З. М., Полетаева Н. Н., Лаврова Н. В., Аксенова Е. И., Семихин А. С., Карягина А. С., Лунин В. Г. Получение и характеристика коллагенсвязывающих доменов из фактора фон Виллебранда человека | Контакты, аннотация, ключевые слова, литература |
31 |
ХРОНИКА |
|
| Лимборская С. А. Международная Школа молодых ученых по молекулярной генетике “Геномика и эволюция” (1—5 декабря 2008 г.) | 35 |
|
Наименование |
Часы |
Вид обучения |
Форма обучения |
Вариант обучения |
|
Актуальные вопросы менеджмента в здравоохранении |
72 |
повышение квалификации |
очная |
аккредитовано в системе НМФО |
|
Базовая сердечно-легочная реанимация с использованием автоматического наружного дефибриллятора» «Basic life support and automated external defibrillation (BLS/AED provider) |
18 |
повышение квалификации |
очно-дистанционная |
аккредитовано в системе НМФО |
|
Базовые основы системы обязательного медицинского страхования |
36 |
повышение квалификации |
дистанционная |
аккредитовано в системе НМФО |
|
Бережливые (Lean) технологии в медицинском учреждении. |
36 |
повышение квалификации |
дистанционная |
аккредитовано в системе НМФО |
|
Вопросы диагностики,лечения и профилактики наследственных болезней |
72 |
повышение квалификации |
очная |
аккредитовано в системе НМФО |
|
Генетика. Избранные вопросы медицинской генетики |
144 |
повышение квалификации |
очная |
традиционное образование |
|
Генетика. Пренатальная диагностика врожденных пороков развития и наследственных болезней |
72 |
повышение квалификации |
очная |
аккредитовано в системе НМФО |
|
Информационные технологии в управлении проектами |
36 |
повышение квалификации |
дистанционная |
аккредитовано в системе НМФО |
|
Информационные технологии для медицинских специалистов |
72 |
повышение квалификации |
очная |
аккредитовано в системе НМФО |
|
Информационные технологии организации проектной деятельности |
18 |
повышение квалификации |
очная |
аккредитовано в системе НМФО |
|
Информационный менеджмент в здравоохранении |
72 |
очная |
очная |
аккредитовано в системе НМФО |
|
Клиническая лабораторная диагностика |
576 |
профессиональная переподготовка |
очная |
традиционное образование |
|
Лабораторная генетика |
504 |
профессиональная переподготовка |
очная |
традиционное образование |
|
Лабораторная генетика |
216 |
повышение квалификации |
очная |
традиционное образование |
|
Медико-социальная экспертиза |
72 |
повышение квалификации |
дистанционная |
аккредитовано в системе НМФО |
|
Медицинская статистика |
144 |
повышение квалификации |
очная |
традиционное образование |
|
Методы статистической обработки результатов медицинских исследований |
108 |
повышение квалификации |
дистанционная |
аккредитовано в системе НМФО |
|
Надлежащая клиническая практика. |
72 |
повышение квалификации |
дистанционная |
аккредитовано в системе НМФО |
|
Наследственные болезни пищеварительной системы |
72 |
повышение квалификации |
очная |
аккредитовано в системе НМФО |
|
Наследственные нарушения обмена соединительной ткани |
72 |
повышение квалификации |
очная |
аккредитовано в системе НМФО |
|
Неотложные состояния в кардиологии |
144 |
повышение квалификации |
очная |
традиционное образование |
|
Неотложные состояния в работе врачей первичного звена здравоохранения |
144 |
повышение квалификации |
дистанционная |
традиционное образование |
|
Оборот наркотических средств, психотропных веществ и их прекурсоров в фармацевтических и медицинских организациях |
72 |
повышение квалификации |
очная |
аккредитовано в системе НМФО |
|
Организация здравоохранения и общественное здоровье |
576 |
профессиональная переподготовка |
дистанционная |
традиционное образование |
|
Организация здравоохранения и общественное здоровье |
504 |
профессиональная переподготовка |
очная |
традиционное образование |
|
Орфанные болезни в практике врачей различных специальностей |
144 |
повышение квалификации |
очная |
традиционное образование |
|
Орфанные болезни в практике врачей различных специальностей |
36 |
повышение квалификации |
очная |
аккредитовано в системе НМФО |
|
Основы классического массажа |
144 |
повышение квалификации |
очная |
традиционное образование |
|
Основы статистического анализа медико-биологических данных |
144 |
повышение квалификации |
очная |
традиционное образование |
|
Планирование и статистический анализ результатов медико-биологических исследований |
216 |
повышение квалификации |
очная |
традиционное образование |
|
Правовые основы оборота наркотических средств, психотропных веществ и их прекурсоров в медицинской практике |
36 |
повышение квалификации |
дистанционная |
аккредитовано в системе НМФО |
|
Пренатальная диагностика врожденных пороков развития и наследственных болезней |
36 |
повышение квалификации |
очная |
аккредитовано в системе НМФО |
|
Применение информационных технологий в медицинской практике |
144 |
повышение квалификации |
очная |
традиционное образование |
|
Применение информационных технологий в медицинской практике и организации здравоохранения |
216 |
повышение квалификации |
очная |
традиционное образование |
|
Психология и педагогика высшей школы |
72 |
повышение квалификации |
очная |
аккредитовано в системе НМФО |
|
Сердечно-легочная реанимация с автоматическим внешним дефибриллятором (СЛР/АВД) |
8 |
повышение квалификации |
очная |
традиционное образование |
|
Сердечно-легочная реанимация с автоматическим внешним дефибриллятором (СЛР/АВД) -Курс Европейского Совета по Реанимации – Basic Life Support / Automated External Defibrillation (ERC/AED provider) |
8 |
повышение квалификации |
очная |
традиционное образование |
|
Симуляция в медицинском образовании (практический курс) |
16 |
повышение квалификации |
очная |
традиционное образование |
|
Статистический учет в медицинской организации |
36 |
повышение квалификации |
очная |
аккредитовано в системе НМФО |
|
Тиреоидология |
108 |
повышение квалификации |
очная |
традиционное образование |
|
Фармакогнозия |
36 |
повышение квалификации |
дистанционная |
традиционное образование |
|
Фармакология |
72 |
повышение квалификации |
очная |
традиционное образование |
|
Фармацевтическая технология |
108 |
повышение квалификации |
очная |
традиционное образование |
|
Фармацевтическая химия |
72 |
повышение квалификации |
очная |
традиционное образование |
|
Фармацетическая химия |
72 |
повышение квалификации |
дистанционная |
традиционное образование |
|
Экпертиза временной нетрудоспособности, медико-социальная экспертиза, контроль качества оказания медицинской помощи |
216 |
повышение квалификации |
дистанционная |
традиционное образование |
|
Экспертиза временной нетрудоспособности |
72 |
повышение квалификации |
дистанционная |
аккредитовано в системе НМФО |
|
Экспертиза нетрудоспособности |
72 |
повышение квалификации |
очная |
аккредитовано в системе НМФО |
|
Экспертиза нетрудоспособности и управление качеством медицинской помощи |
72 |
повышение квалификации |
очная |
аккредитовано в системе НМФО |
 Базовый уровень (уровень А)
Базовый уровень (уровень А) Основы доказательной медицины и методологии планирования клинических исследований
Основы доказательной медицины и методологии планирования клинических исследованийИнвитро.
 Генетические предрасположенности, узнать цены на анализы и сдать в Москве Тромбозы: расширенная панель
Генетические предрасположенности, узнать цены на анализы и сдать в Москве Тромбозы: расширенная панельВыявление индивидуальных особенностей в 6 генах системы гемостаза для оценки наличия факторов риска развития тромбоза и повышения уровня гомоцистеина (гены протромбина, фактора Лейдена и ферментов реакций фолатного цикла).
ГипергомоцистеинемияВыявление изменений в основных генах ферментов фолатного цикла для оценки наличия склонности к гипергомоцистеинемии (рекомендовано оценивать в комплексе с иммунохимическим тестом на определение уровня гомоцистеина).
Гиперагрегация тромбоцитов, полиморфизм гена ITGA2 с.759 С>TИсследование полиморфизмов в генах интегрина альфа-2 и тромбоцитарного гликопротеина 1b проводят для выявления генетической предрасположенности к раннему развитию инфаркта миокарда, ишемического инсульта, тромбоэмболии, а также для оценки риска развития тромбозов. На бланке результата выдается информация о полиморфизмах, полученная при молекулярно-генетическом исследовании, с комментариями.
Определение полиморфизмов в гене тромбоцитарного рецептора фибриногена (β3-интегрина) выполняют для выявления наследственной предрасположенности к тромбофилическим состояниям. На бланке результата выдается информация о полиморфизмах, полученная при молекулярно-генетическом исследовании, с комментариями.
Сердечно-сосудистые заболеванияВ процессе исследования выявляют генетические факторы риска развития артериальной гипертензии, атеросклероза, ишемической болезни сердца, инфаркта миокарда, ишемического инсульта.
Артериальная гипертензия (полная панель)Анализ полиморфизмов в генах ACE, AGT, NOS3 дает возможность обнаружить наследственные факторы риска развития артериальной гипертензии. На бланке результата выдается информация о полиморфизмах, полученная при молекулярно-генетическом исследовании, с комментариями.
Артериальная гипертензия, связанная с нарушениями в ренин-ангиотензиновой системеТест позволяет определить наличие генетических факторов риска развития артериальной гипертензии в результате сужения просвета сосудов и нарушения водно-солевого баланса, возникающих при наличии полиморфизмов в генах ACE, AGT. На бланке результата выдается информация о полиморфизмах, полученная при молекулярно-генетическом исследовании, с комментариями.
На бланке результата выдается информация о полиморфизмах, полученная при молекулярно-генетическом исследовании, с комментариями.
Тест используют при диагностике болезни Крона, для определения прогноза тяжести течения заболевания и риска развития осложнений. Также исследование применяют для дифференциальной диагностики болезни Крона с язвенным колитом и в качестве прогностического теста у родственников пациентов с болезнью Крона.
Поражения печени, гены ATP7B, PNPLA3, SERPINA1, ч.м.Исследование используется для дифференциальной диагностики генетических причин патологии печени – дефицита альфа-1-антитрипсина, болезни Вильсона-Коновалова, классического гемохроматоза и злокачественной формы неалкогольной жировой болезни печени
Анализ перестроек 1 хромосомы (FISH, колич.)Исследование показано для диагностики, определения прогноза течения заболевания, подбора адекватной терапии и мониторинга минимальной остаточной болезни пациентов с множественной миеломой.
Синдром множественной эндокринной неоплазии 2В типа относится к группе семейных опухолевых синдромов, ассоциированных со специфическими мутациями протоонкогена RET, которые выявляют в процессе исследования.
Синдром Жильбера, UGT1A1Генетическая диагностика синдрома Жильбера – неконъюгированной доброкачественной гипербилирубинемии – основана на исследовании возможных мутаций в промоторной области гена UGT1A1.
Остеопороз: полная панельИсследование проводят при наличии нарушений минерального обмена, а также при отягощенном семейном анамнезе по заболеваниям костной системы.
Остеопороз: сокращённая панельИсследование проводят при наличии нарушений минерального обмена, а также при отягощенном семейном анамнезе по заболеваниям костной системы.
Остеопороз: рецептор витамина DИсследование генетических факторов риска развития остеопороза проводят при отягощенном семейном анамнезе по заболеваниям костного аппарата, а также при наличии нарушений минерального обмена. На бланке результата выдается информация о полиморфизмах, полученная при молекулярно-генетическом исследовании, с комментариями.
На бланке результата выдается информация о полиморфизмах, полученная при молекулярно-генетическом исследовании, с комментариями.
Выявление индивидуальных особенностей в основных генах ферментов фолатного цикла для оценки наличия склонности к гипергомоцистеинемии (рекомендовано оценивать в комплексе с иммунохимическим тестом на определение уровня гомоцистеина).
Наследственный гемохроматоз, I тип. HFEВыявление 2 наиболее часто встречаемых мутаций в гене HFE для оценки риска развития гемохроматоза 1-го типа. Рекомендовано при выявлении повышения концентрации ферритина и % насыщения трансферрина железом в сыворотке крови.
Описание результатов генетического теста 2 категории сложности (№№ 118ГП/БЗ, 121ГП/БЗ, 123ГП/БЗ, 131ГП/БЗ, 141ГП/БЗ, 149ГП/БЗ, 150ГП/БЗ, 115ГП/БЗ, 152ГП/БЗ, 124ГП/БЗ, 154ГП/БЗ)Исследование включает описание врачом-генетиком результатов генетических анализов, которые относятся ко второй категории сложности.
Исследование включает описание врачом-генетиком результатов генетических анализов, которые относятся к третьей категории сложности.
Ингибиторы АПФ, флувастатин, блокаторы рецепторов АТIIАнализ полиморфизмов в гене ACE необходим для прогнозирования нефропротективного эффекта ингибиторов ангиотензин-превращающего фермента (АПФ) – физиологического регулятора артериального давления и водно-солевого обмена при недиабетических заболеваниях. Посредством исследования можно определить генетические маркеры эффективности атенолола при артериальной гипертензии с гипертрофией левого желудочка или флувастатина при ишемической болезни сердца.
Лозартан/ирбесартанВ процессе исследования проводится анализ полиморфизмов в гене CYP2C9, который выступает генетическим маркером риска нарушений метаболизма блокаторов рецепторов ангиотензина II.
Выявление изменений в основных генах ферментов фолатного цикла для оценки вероятности развития побочных реакций при приеме метотрексата.
Нестероидные противовоспалительные препаратыАнализ полиморфизмов в гене CYP2C9 выполняют с целью выявления наследственных факторов развития побочных реакций по типу желудочных кровотечений при приеме нестероидных противовоспалительных препаратов (НПВП).
Бета-адреноблокаторы. Ген CYP2D6. Фармакогенетика.Цитохром CYP2D6 участвует в метаболизме лекарственных препаратов (β-адреноблокаторов, антиаритмиков, аналептиков, антидепрессантов и наркотических анальгетиков), применяемых при лечении ряда сердечно-сосудистых заболеваний и психических расстройств. Исследование полиморфизмов в гене CYP2D6 позволяет выявить людей со сниженной активностью CYP2D6, поскольку таким пациентам необходимо индивидуально подбирать более низкие дозы препаратов.
Цитохром СYP2D6: ген СYP2D6Тест целесообразно проводить перед плановым назначением антигипертензивных, антиаритмических, психотропных препаратов, а также при длительном их приеме.
Анализ наличия полиморфизмов в гене цитохрома Р450 проводят для выявления наследственных факторов нарушения детоксикации. CYP2C9 участвует в метаболизме лекарственных средств. При снижении активности цитохрома CYP2C9 метаболизм препаратов замедляется, в результате чего происходит увеличение их концентрации в крови, что может быть причиной развития нежелательных реакций.
ДНК-тест для Ивана Грозного. Как генетика открывает тайны жизни и истории | ОБЩЕСТВО:Люди | ОБЩЕСТВО
На вопросы «АиФ на Дону» о загадках генетики ответил доктор биологических наук, заведующий лабораторией «Идентификация объектов биологического происхождения» Академии биологии и биотехнологии Южного федерального университета Игорь Корниенко.
История в лаборатории
Юлия Морозова, «АиФ на Дону»: Вы занимаетесь именно молекулярной генетикой, что это такое?
Игорь Корниенко. Родился в Ростове-на-Дону 14 декабря 1970 г.
Родился в Ростове-на-Дону 14 декабря 1970 г.
В 1994 году окончил Ростовский государственный университет, биолого-почвенный факультет.
В 2000-м окончил химический факультет ЮФУ (РГУ).
В 1997 г. защитил кандидатскую диссертацию, в 2005-м докторскую диссертацию.
Автор более 160 научных работ.
Игорь Корниенко: Классическая генетика (селекция) выводит новые и улучшает старые сорта растений или породы животных. Селекционера интересуют признаки, полезные для человека, и его не интересует, почему, например, горошек зелёный. А молекулярная генетика разбирается с тем, какой ген отвечает за этот цвет.
Вообще генетика, на мой взгляд, одна из самых интересных наук. Представьте: человек состоит из десятков триллионов клеток, каждая – это самостоятельный биохимический котёл, в котором протекают разные сложные процессы. В клетке есть компактно упакованные молекулы ДНК. Если их развернуть, то длина нитей ДНК одной клетки достигнет двух метров. Умножим это на десятки триллионов клеток и получим огромную цепочку ДНК, которой можно обернуть не только Землю, но и всю Солнечную систему.
– И эту цепочку вы изучаете?
– Основная моя работа – это исследование ДНК древних животных и людей. Несколько лет назад московские учёные вскрыли два саркофага. Историки предполагали, что там покоятся родственники Ивана Грозного по отцовской линии. Нам на исследование предоставили найденные останки.
Сложность заключалась в том, что наши коллеги дали нам лишь самую общую информацию о присланных образцах. Мы поработали с ДНК и определили, что это останки матери и дочери. Причём мать умерла при рождении дочери. Позже удалось установить, что это дочь (Евдокия) и внучка (Анастасия) Софии Палеолог, которая приходилась бабушкой Ивану Грозному. Так генетика помогла историкам.
Мы также тесно сотрудничаем с археологами. Работали с захоронениями I века нашей эры в античном городе Танаисе. Определяли, как влияют различные типы почв юга России на сохранность ДНК в древних костях.
Кстати, на Дону нам попадались даже африканские кости. Недавно в нашей лаборатории совместно с Донской митрополией мы проводили генетическую экспертизу останков святого Павла Таганрогского. По поводу этого святого были различные спекуляции и слухи, будто бы это останки императора Александра I. Несмотря на плохую сохранность ДНК, мы доказали, что мощи принадлежат одному человеку, Павлу Таганрогскому.
По поводу этого святого были различные спекуляции и слухи, будто бы это останки императора Александра I. Несмотря на плохую сохранность ДНК, мы доказали, что мощи принадлежат одному человеку, Павлу Таганрогскому.
– Несколько лет назад на острове Малый Ляховский (Новосибирский архипелаг в море Лаптевых) была обнаружена хорошо сохранившаяся туша мамонта. А можно ли клонировать это древнее животное?
– Действительно, состояние найденной 60-летней самки шерстистого мамонта (она погибла примерно 30-40 тысяч лет назад) было хорошее. Мышечные волокна хобота сохранили свою структуру и имели красный цвет. Сейчас совместно с красноярскими и японскими коллегами мы проводим исследование частей этой туши. Но не на предмет создания клона. Ведь даже при клонировании овечки Долли использовали 227 яйцеклеток, чтобы получился один жизнеспособный клон. Остальные попытки были неудачны. То есть процент удачи был менее 0,5.
Но в случае с Долли брали живые яйцеклетки и ядра клеток. А у замороженной самки мамонта нет целых клеток и ядер, всё повреждено кристаллами льда. Давайте представим на минуту, что нам каким-то образом удалось добыть целые ядра клеток шерстистого мамонта. Но для его клонирования нам нужны десятки яйцеклеток азиатского слона. Их беременность длится два года, ждать положительные результаты придётся долго. А представьте стоимость этого клонированного мамонтёнка! По моим оценкам, вероятность клонирования таким способом шерстистого мамонта близка к нулю.
А у замороженной самки мамонта нет целых клеток и ядер, всё повреждено кристаллами льда. Давайте представим на минуту, что нам каким-то образом удалось добыть целые ядра клеток шерстистого мамонта. Но для его клонирования нам нужны десятки яйцеклеток азиатского слона. Их беременность длится два года, ждать положительные результаты придётся долго. А представьте стоимость этого клонированного мамонтёнка! По моим оценкам, вероятность клонирования таким способом шерстистого мамонта близка к нулю.
Но есть более перспективный метод – это редактирование генома азиатского (индийского) слона: постепенное внесение изменений в его ДНК. С каждым новым поколением родившиеся слонята будут всё более походить на мамонтят. Это долго и потребует огромных финансовых затрат, но более реально.
Рай для учёного
– Игорь Валерьевич, вы через несколько дней летите в Японию. Что именно интересует ваших зарубежных коллег?
– Делимся опытом по изучению древней ДНК. Кроме того, у нас с этим университетом совместный проект по исследованию донских захоронений хазар, датируемых VIII-IX веками до нашей эры. Сейчас хазарский каганат – это почва для лженаучных открытий, идут постоянные споры о том, к каким народам они близки.
Кроме того, у нас с этим университетом совместный проект по исследованию донских захоронений хазар, датируемых VIII-IX веками до нашей эры. Сейчас хазарский каганат – это почва для лженаучных открытий, идут постоянные споры о том, к каким народам они близки.
Зарубежным генетикам Россия всегда была интересна, особенно Кавказ. Возьмём, например, США. Там выделяются всего несколько групп: европеоиды – к ним относятся все представители белой расы, монголоиды (джапаниз), коренные американцы (племена индейцев), латиноамериканцы и афроамериканцы. Всё обобщено и размыто. А возьмём, например, Дагестан. Здесь представлены более 30 народностей (аварцы, лезгины, кумыки и т.д.), они досконально прослеживают свои корни, свою историю, а кроме того, имеют чёткие различительные национальные признаки.
Но для любых исследований нам необходимы огромные мощности. Работать с древними костями тяжело. После смерти ДНК подвергается необратимым изменениям разложению от воздействия жары, влажности, ультрафиолетового излучения. Даже если вы получили генетический профиль, то нет никакой гарантии, что он не изменился под действием внешних факторов, а такой и был тысячи лет назад.
Даже если вы получили генетический профиль, то нет никакой гарантии, что он не изменился под действием внешних факторов, а такой и был тысячи лет назад.
Поэтому для исследования берём одновременно несколько костных фрагментов от одного скелета. Чтобы обобщить и анализировать огромное количество материала, нужны мощные компьютеры, которые нам предоставляют специалисты Окинавского университета. Вот такое взаимовыгодное сотрудничество.
– Расскажите подробнее об уникальном исследовании, благодаря которому военнослужащий, пропавший без вести во время Второй мировой войны, обрёл имя и родственников.
– Бои на линии обороны «Миус-Фронта» шли ожесточённые, на некоторых участках потери нашей армии составили до трёх человек на один квадратный метр. Сейчас здесь постоянно работают патриотические поисковые отряды.
Но вот установить личность найденных красноармейцев очень трудно: медальоны либо пустые (солдаты из-за суеверия не вкладывали в них бумажку со своими данными), либо документ в капсулах испорчен влагой.
В январе 2016 года в районе Самбека обнаружили обломки самолёта, рядом – скелеты трёх человек. Серийный номер мотора уцелел, и поисковики связались с архивами. По документам стало известно, что этот самолёт пропал без вести 17 октября 1941 года. Удалось также узнать фамилии членов экипажа, среди них был стрелок люковой установки, младший сержант Владимир Дзюба. Но как определить, кто есть кто?
На помощь пришёл молекулярно-генетический анализ. Этот метод сравнительный, нужен биологический материал погибшего и его прямых родственников. Мы стали разыскивать потомков бойцов, и нам улыбнулась удача: в Краснодаре нашлась родная племянница красноармейца Дзюбы. Это помогло произвести все необходимые исследования. Через 75 лет после войны мы смогли вернуть погибшему сержанту имя и семью.
Было бы хорошо создать сейчас генетическую базу данных (ДНК-депозитарий) родственников пропавших без вести красноармейцев. Найдут, например, через 10 лет останки, а сравнивать материал будет уже не с кем, прямых родственников становится всё меньше. И солдата ждёт безымянная могила.
И солдата ждёт безымянная могила.
Счётчик жизни
– В ДНК содержится так много информации, а, может быть, там «прописана» и продолжительность нашей жизни?
– Да, это можно узнать. Существует лимит Хейфлика, то есть клетка делится примерно 50-60 раз, а потом умирает. Этот процесс – хотите, назовите его старением – начинается уже у ребёнка во чреве матери. Поэтому с биологической точки зрения не совсем правильно считать днём его рождения дату появления на свет.
На концах нашего ДНК есть «колпачки», они называются теломеры. При каждом клеточном делении размер молекул ДНК немного уменьшается. Рано или поздно теломеры сократятся до критической длины.
И вот тогда запускается каскад болезней старости (онкология, сердечная, лёгочная недостаточность), снижается уровень иммунитета. Такой молекулярный «счётчик» у кого-то может щёлкнуть в 100 лет, а у кого-то в 70.
При рождении двух младенцев можно сделать вероятностный прогноз, кто из них сколько может прожить. Однако не стоит забывать, что не всё определяется нашим геномом, среда тоже оказывает огромное влияние на продолжительность жизни. Нобелевским лауреатом Элизабет Блэкберн было показано, что курение укорачивает теломеры.
Однако не стоит забывать, что не всё определяется нашим геномом, среда тоже оказывает огромное влияние на продолжительность жизни. Нобелевским лауреатом Элизабет Блэкберн было показано, что курение укорачивает теломеры.
Поэтому не всё зависит от того, какую ДНК вы получили в наследство от родителей. Правильно питайтесь, занимайтесь спортом, и долгая и счастливая жизнь будет более вероятна.
Классическая генетика – обзор
1 Введение
Классическая генетика заложила основы понимания функции генов. Традиционно анализируемые мутации были либо спонтанными, либо вызванными тем, что мы сегодня назвали бы «генотоксическим стрессом». Тщательный анализ полученных фенотипов, их паттернов наследования и выводы из объединения разных генотипов путем селекции или других «традиционных» методов перетасовки геномов позволили глубоко проникнуть во многие биологические процессы, даже не имея возможности определить молекулярную природу мутации.Большинство мутантов имели либо явную потерю функции, либо усиление функции в результате точечных мутаций, делеций, инверсий или транслокаций. Однако некоторые из этих мутаций привели к появлению гипоморфов, то есть аллелей, характеризующихся частичной потерей функции. Некоторые из них были условными в том смысле, что экспрессивность мутантного аллеля могла контролироваться внешними сигналами. Хорошо известными примерами являются аллели ts , в которых мутация делает функциональность соответствующего белка чувствительной к температуре.Возможность произвольно контролировать фенотип данного генотипа в значительной степени способствовала пониманию физиологических процессов и процессов развития, в частности, в микробной генетике и в генетике Drosophila . Генетика мышей не могла обеспечить сравнимую глубину понимания функции генов до появления генной инженерии. Однако возможность создания трансгенных мышей путем случайной интеграции трансгенов и последующее развитие технологии эмбриональных стволовых клеток (ES-клетки) с ее потенциалом вносить целевые мутации в геном сделали мышь пригодной для генетических подходов на молекулярном уровне.
Однако некоторые из этих мутаций привели к появлению гипоморфов, то есть аллелей, характеризующихся частичной потерей функции. Некоторые из них были условными в том смысле, что экспрессивность мутантного аллеля могла контролироваться внешними сигналами. Хорошо известными примерами являются аллели ts , в которых мутация делает функциональность соответствующего белка чувствительной к температуре.Возможность произвольно контролировать фенотип данного генотипа в значительной степени способствовала пониманию физиологических процессов и процессов развития, в частности, в микробной генетике и в генетике Drosophila . Генетика мышей не могла обеспечить сравнимую глубину понимания функции генов до появления генной инженерии. Однако возможность создания трансгенных мышей путем случайной интеграции трансгенов и последующее развитие технологии эмбриональных стволовых клеток (ES-клетки) с ее потенциалом вносить целевые мутации в геном сделали мышь пригодной для генетических подходов на молекулярном уровне. и, таким образом, к наиболее подходящей модельной системе млекопитающих.Во многих модельных системах «классической» генетики возможность создания условных мутантов значительно расширила спектр вопросов, которые можно задать, особенно при введении потенциально вредных мутаций. У мышей обусловленность может быть достигнута за счет пространственного и временного контроля над рекомбиназами (Anastassiadis et al. , 2010), что обычно приводит к необратимым генетическим изменениям. При наличии подходящих векторов таргетинга этот подход может гарантировать полный нулевой генотип.Напротив, истинная обусловленность, то есть обратимое изменение фенотипа, может быть достигнута системами экспрессии, контролирующими активность генов на уровне транскрипции. В основном это гетерологичные бинарные системы экспрессии, состоящие из сконструированного фактора транскрипции и соответствующей ответной единицы, включающей интересующий трансген (Gossen et al. , 1993). Было разработано несколько таких систем, в которых активность генов можно контролировать на уровне транскрипции, которые функционируют в культивируемых клетках различного происхождения.Однако лишь некоторые из них широко применяются. Разнообразие транскрипционно контролируемых систем экспрессии с установленной функциональностью у мышей еще более ограничено и почти исключительно ограничивается системами Tet-On и Tet-Off.
и, таким образом, к наиболее подходящей модельной системе млекопитающих.Во многих модельных системах «классической» генетики возможность создания условных мутантов значительно расширила спектр вопросов, которые можно задать, особенно при введении потенциально вредных мутаций. У мышей обусловленность может быть достигнута за счет пространственного и временного контроля над рекомбиназами (Anastassiadis et al. , 2010), что обычно приводит к необратимым генетическим изменениям. При наличии подходящих векторов таргетинга этот подход может гарантировать полный нулевой генотип.Напротив, истинная обусловленность, то есть обратимое изменение фенотипа, может быть достигнута системами экспрессии, контролирующими активность генов на уровне транскрипции. В основном это гетерологичные бинарные системы экспрессии, состоящие из сконструированного фактора транскрипции и соответствующей ответной единицы, включающей интересующий трансген (Gossen et al. , 1993). Было разработано несколько таких систем, в которых активность генов можно контролировать на уровне транскрипции, которые функционируют в культивируемых клетках различного происхождения.Однако лишь некоторые из них широко применяются. Разнообразие транскрипционно контролируемых систем экспрессии с установленной функциональностью у мышей еще более ограничено и почти исключительно ограничивается системами Tet-On и Tet-Off.
Молекулярная генетика (Стэнфордская энциклопедия философии)
Термин молекулярная генетика теперь излишен, потому что современная генетика полностью молекулярна. Генетика не придумана двух наук, молекулярной и немолекулярной.Тем не менее, практикующие биологи до сих пор используют этот термин. Когда они это сделают, они обычно относится к набору лабораторных методов, направленных на идентификация и / или манипулирование сегментами ДНК, участвующими в синтезе важных биологических молекул. Ученые часто говорят и пишут о применении этих методов в широком спектре биомедицинские науки. Для них молекулярная генетика – это следственный подход , который предполагает применение лабораторные методы и стратегии исследования.Такой подход предполагает базовые знания об экспрессии и регуляции генов на молекулярный уровень.
Философский интерес к молекулярной генетике, однако, был сосредоточен, а не на исследовательские подходы или лабораторные методы, но теоретически. Рано философское исследование касалось базовой теории о состав, экспрессия и регуляция генов. Наибольшее внимание сосредоточен на проблеме теоретического редукционизма. Мотивация Вопрос касался ли классическая генетика, наука Т.ЧАС. Моргана и его сотрудников сводили к молекулярной генетике. С ростом генетики развития и биологии развития, философское внимание впоследствии сместилось в сторону критики фундаментальная теория, связанная с современной генетикой. В фундаментальная теория касается не только макияжа, экспрессия и регуляция генов, но также и общая роль генов внутри организма. Согласно фундаментальной теории, гены и ДНК направляет все жизненные процессы, предоставляя информацию, которая определяет развитие и функционирование организмов.
Эта статья начинается с краткого обзора основной теории. связаны с молекулярной генетикой. Поскольку эта теория включает идеи из школы классической генетики Моргана, полезно набросайте его развитие от Моргана генетика. Изучив основную теорию, я исследую четыре вопроса. ведущие философские исследования молекулярной генетики. Первое Вопрос спрашивает, была ли классическая генетика сведена или будет сведена к молекулярная генетика. Второй вопрос касается концепции гена и изжила ли она свою полезность.Третий вопрос касается жизнеспособность фундаментальной теории. Четвертый вопрос, который еще не привлекла к себе особого философского внимания, спрашивает, почему так много биологических исследований сосредоточено на генах и ДНК.
2.1 Базовая теория классической генетики
Основная теория, связанная с классической генетикой, предоставила объяснения передачи признаков от родителей к потомству. Морган и его сотрудники провели концептуальное разделение между генетический состав организма, названный его генотипом , и его наблюдаемое проявление назвало его фенотип (см. вход на различие генотипа / фенотипа).Связь между ними рассматривалась как причинная: генотип в соединение с окружающей средой производит фенотип. Теория объяснила передача фенотипических различий от родителей к потомству посредством после передачи генных различий от поколения к генерации и приписывая наличие альтернативных черт наличие альтернативных форм генов.
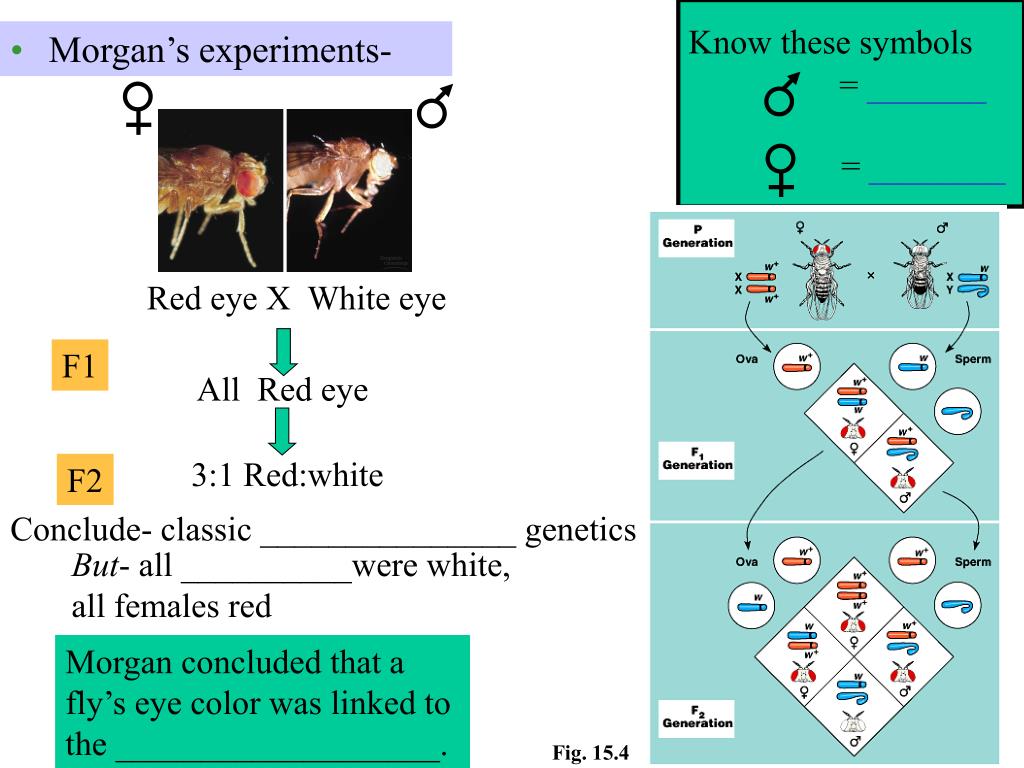
Я проиллюстрирую классический способ объяснительного рассуждения с помощью простой исторический пример с плодовой мушкой Drosophila Меланогастор .Стоит подчеркнуть, что способ рассуждения иллюстрированный этим историческим примером, по-прежнему является важным способом рассуждения в генетике сегодня, в том числе то, что иногда называют молекулярная генетика.
Гены Drosophila попарно расположены в соответствующих позиции на четырех парах хромосом, содержащихся в каждой клетке мухи. Мутант по цвету глаз, известный как пурпурный, связан с с геном, расположенным на хромосоме II. Две копии этого гена, существующие либо в мутированной, либо в нормальной форме дикого типа, расположены в тот же локус (соответствующая позиция) в двух вторая хромосома.Альтернативные формы гена, встречающиеся в локусом называются аллели . Передача генов от Отцовство от потомства осуществляется в особом процессе клеточного деление, называемое , мейоз , которое производит клетки гамет содержащие по одной хромосоме из каждого парного набора. Половина набора хромосомы из яйцеклетки и половину набора из сперматозоидов объединяются во время оплодотворение, которое дает каждому потомству копию одного гена от каждого пара генов его родительской женщины и копия одного гена из каждого гена пара своего родителя-мужчины.
Объяснения передачи признаков связаны с наличием альтернативные гены (генотип) наличию альтернативных наблюдаемых черты (фенотип). Иногда это делается с точки зрения доминантные / рецессивные отношения. Например, фиолетовый цвет глаз – это рецессивный к символу дикого типа (красный цвет глаз). Это означает летает с двумя копиями пурпурного аллеля (мутантная форма ген, который обозначен пр ) имеют пурпурные глаза, но гетерозигот , летает с одной копией пурпурного аллеля и одна копия аллеля дикого типа (обозначена +), имеет нормальный дикий тип глаза (как и мухи с двумя копиями аллеля дикого типа).См. Таблицу 1.
Чтобы увидеть, как классическая теория объясняет передачу признаков, рассмотрим крест красноглазых самок с пурпурглазыми самцами, выполненный сотрудниками Моргана. Потомство у всех были красные глаза. Так черта красных глаз передалась от самок. всему их потомству, даже если потомство у родителей-мужчин были пурпурные глаза. Классическое объяснение этого паттерн наследования продолжается, как и все классические объяснения паттерны наследования в два этапа.
Первый этап отвечает за передачу генов и выглядит следующим образом: следует (рисунок 1): каждое потомство получило по одной копии хромосомы II от каждого родителя. Хромосомы материнского происхождения должны иметь содержал аллель дикого типа (поскольку обе вторые хромосомы каждого родительская женщина, использованная в эксперименте, содержала аллель дикого типа – это было известно на основании предыдущих экспериментов). По отцовской производные хромосомы должны содержать аллель пурпурного цвета (поскольку оба Вторая хромосома каждого родителя мужского пола содержала аллель пурпурного цвета – это было выведено из знания, что фиолетовый рецессивен к красному цвет глаз).Следовательно, все потомки были гетерозиготными ( пр / + ). Объяснив генетический состав потомства отслеживая передачу генов от родителей к потомству, мы можем приступаем ко второму этапу объяснения: составление вывода о фенотипических проявлениях. Поскольку все потомки были гетерозиготными ( пр / + ), и поскольку фиолетовый рецессивен к дикому типу, у всех потомков был красный цвет глаз (характер дикого типа). См. Рисунок 1.
Обратите внимание, что рассуждения здесь не зависят от идентификации материальный состав, способ действия или общая функция нижележащего ген.Это зависит только от представлений о том, что копии гена распределяется из поколения в поколение и что разница в ген (т.е. разница между пр и + ), Какова бы ни была эта разница, она вызывает фенотипическое различие. Идея необходимо указать, что ген является фактором, создающим разницу: различия в генах вызывают фенотипические различия, в частности генетический и экологический контексты. Эта идея настолько важна, что часто упускают из виду, что он заслуживает артикуляции в качестве принципа (Waters 1994):
Принцип различия : различия в классической генной причине однородные фенотипические различия в конкретных генетических и экологических контексты.
Также стоит отметить, что принцип разности обеспечивает средства для объяснения передачи фенотипических характеристик от от поколения к поколению, не объясняя, как эти характеристики производятся в процессе жизнедеятельности организма разработка. Это эффективно позволило классическим генетикам разработать наука о наследственности без ответов на вопросы о разработка.
Практика классической генетики включала теоретический анализ сложные схемы передачи, включающие рекомбинацию фенотипические признаки.Анализ этих закономерностей дал информацию о основные биологические процессы, такие как хромосомная механика, а также информация о линейном расположении генов в группах сцепления. Эти теоретические объяснения не зависели от представлений о том, какие гены как реплицируются гены, что делают гены или как различия в гены вызывают различия в фенотипических признаках.
2.2 Ответы на молекулярном уровне на вопросы, оставленные классической генетикой
Исследования в области молекулярной биологии и генетики дали ответы на основные вопросы, оставленные без ответа классической генетикой о состав генов, механизм репликации генов, то, что гены делают, и способ, которым различия генов вызывают фенотипические различия.Эти ответы сформулированы в терминах явлений молекулярного уровня, и они обеспечивают большую часть базовой теории , связанной с молекулярными генетика.
Что такое ген? Этот вопрос решен. подробнее в разделе 4 этой статьи, но быстрый ответ для настоящих целей достаточно: гены представляют собой линейные последовательности нуклеотиды в молекулах ДНК. Каждая молекула ДНК состоит из двойного цепь нуклеотидов. В ДНК есть четыре типа нуклеотидов: гуанин, цитозин, тимин и аденин.Пара нуклеотидных цепей в молекуле ДНК закручиваются друг вокруг друга в виде двойного спираль. Две цепи спирали связаны водородными связями между нуклеотиды из соседних цепей. Водородная связь специфична, поэтому что гуанин в одной цепи всегда находится рядом с цитозином в соседняя цепочка (и наоборот) и тимин в одной цепочке всегда расположен рядом с аденином (и наоборот). Следовательно, линейная последовательность нуклеотидов в одной цепи нуклеотидов в молекуле ДНК комплементарен линейной последовательности нуклеотидов в другой цепи молекулы ДНК.Ген – это сегмент нуклеотидов в одном из цепи молекулы ДНК. Конечно, не каждая строка нуклеотиды в ДНК – это ген; сегменты ДНК идентифицируются как гены согласно тому, что они делают (см. ниже).
Как реплицируются гены? Идея о том, что гены сегментов двойной спирали ДНК дает прямой ответ на этот вопрос. Гены точно воспроизводятся, когда парные цепи молекулы ДНК раскручиваются, и по бокам образуются новые цепи. разделение цепей спариванием комплементарных нуклеотидов.Когда процесс завершен, две копии оригинальной двойной спирали имеют были сформированы и, следовательно, гены в исходной молекуле ДНК были эффективно тиражируется.
Что делают гены? Грубо говоря, гены служат матрицы в синтезе молекул РНК. В результате линейная последовательность нуклеотидов во вновь синтезированной молекуле РНК соответствует линейной последовательности нуклеотидов в сегменте ДНК используется в качестве шаблона. Различные молекулы РНК выполняют разные функции. роли в клетке, и многие молекулы РНК играют роль матрицы в синтез полипептидных молекул.Недавно синтезированные полипептиды представляют собой линейные последовательности аминокислот, которые составляют белки и белки играют в клетке самые разные функциональные роли и организм (и окружающая среда). Способность полипептида функционировать в конкретные пути зависят от линейной последовательности аминокислот, из которых он сформирован. И эта линейная последовательность соответствует линейной последовательности триплетов нуклеотидов в РНК (кодонов), что, в свою очередь, соответствует линейной последовательности нуклеотидов в сегментах ДНК, и это последний сегмент является геном этого полипептида.
Как различия в генах вызывают различия в фенотипические признаки? Скромный ответ, данный выше на вопрос “Что делают гены?” дает основу для объяснения того, как различия в генах могут приводить к о различиях фенотипических признаков. Разница в нуклеотиде последовательность гена приведет к различию в нуклеотиде последовательность молекул РНК, что, в свою очередь, может привести к различию в аминокислотная последовательность полипептида. Различия в линейном последовательности аминокислот в полипептидах (и в линейных последовательностях нуклеотидов в функциональных молекулах РНК) могут влиять на роли, которые они играть в клетке и организме, иногда оказывая наблюдается как фенотипическое различие.Мутации (различия в гены), идентифицированные группой Моргана (например, пурпурная мутация), имеют обычно идентифицируются как различия в нуклеотидных последовательностях в ДНК.
2.3 Различие между основными и фундаментальными теориями молекулярной генетики
Скромный ответ на вопрос “Что делают гены?” что они «кодируют» или «определяют» линейный последовательности в молекулах РНК и полипептидах, синтезированных в клетка. (Даже этот скромный ответ требует уточнения, поскольку РНК молекулы часто соединяются и редактируются способами, влияющими на линейный последовательность аминокислот в конечном продукте полипептида.) Но Биологи также предложили гораздо менее скромный ответ. Смелее Ответ – часть обширной фундаментальной теории. Согласно этому теории, гены – это «фундаментальные» объекты, которые «Направлять» развитие и функционирование организмов посредством «Производящие» белки, которые, в свою очередь, регулируют все важные клеточные процессы. Часто утверждают, что гены обеспечивают «Информация», «план» или «Программа» для организма. Полезно различать эта радикальная фундаментальная теория о якобы фундаментальная роль генов из скромных, основных теория о том, что гены делают в отношении синтеза РНК и полипептиды.
Философов науки заинтриговали идеалы редукционизм и грандиозный план, согласно которому вся наука однажды станет «Сводится» к универсальной науке о фундаментальная физика (см. запись на межтеоретические отношения в физике) для философских и научных концепций редукционизма в в контексте физической науки). Философские редукционисты считают, что научное знание прогрессирует, когда наука более высокого уровня (например, химия) сводятся к наукам более низкого уровня (е.г., физика). Так называемый получил признание научных знания , систематизированные в Nagel (1961) и Hempel (1966), продвигали редукционизм как центральный идеал науки и уверенно утверждал такой большой прогресс был достигнут в сокращении химии до физика. Нагель построил формальную модель редукции и применил ее. чтобы осветить, как наука термодинамика, которая была сформулирована с точки зрения понятий более высокого уровня, таких как давление и температура, было якобы сводится к статистической механике, сформулированной в терминах низкоуровневые концепции ньютоновской динамики, такие как сила и среднее кинетическая энергия.В 1969 году Шаффнер утверждал, что такая же прогресс в генетике, и что наука о классическая генетика сводилась к зарождающейся науке о «Молекулярная генетика». Однако заявление Шаффнера было быстро оспаривается Халлом. Другие философы биологии разработали Антиредукционистские аргументы Халла и почти единодушное мнение о том, что классическая генетика не была и не будет сводиться к молекулярной генетика. Хотя философские доводы в пользу антиредукционизма имеют подверглись сомнению, многие философы все еще полагают, что антиредукционистский взгляд на генетику служит примером антиредукционистский анализ других наук.
3.1 Тезис Шаффнера о том, что классическая генетика теоретически редуцируется
Редукционизм имеет много значений. Например, фраза генетический редукционизм касается идеи, что все биологические явления вызвано генами, и, следовательно, предполагает онтологический смысл редукционизм, согласно которому один вид микро-сущностей (в данном случае ген) исключительно вызывает множество явлений более высокого уровня (в этом случай, биологические особенности, культурные феномены и т. д.).Но не в этом смысл редукционизма, о котором идет речь в философской литература о редукции классической генетики. Эта литература больше занимается эпистемологией, чем метафизикой. Концепция чего-либо редукционизм – это концепция Нагеля о теоретическая редукция . (См. Sarkar 1992 и Schaffner 1993 для обсуждение альтернативных концепций редукции.) Концепция Нагеля, сведение одной науки к другая наука влечет за собой редукцию центральной теории одна наука к центральной теории другой.Нагель верил что такая теоретическая редукция привела к прогрессивным изменениям в научные знания. Он сформулировал два формальных требования к теоретические сокращения.
Первым формальным требованием Нагеля было то, что «законы» редуцированная теория должна быть выведена из законов и связанных с ними согласование определений редукционной теории. Эта выводимость требование было направлено на то, чтобы уловить идею о том, что пояснительное принципы (или законы) теории восстановления должны объяснять объяснительные принципы (или законы) редуцированной теории.Нагеля Второе формальное требование, требование возможности подключения, заключалось в том, что все существенные термины редуцированной теории должны либо содержаться в пределах или быть соответствующим образом связанными с условиями сокращающего теория в виде дополнительных предположений. Возможность подключения требование предполагается из требования выводимости, но делая это явным, помогает подчеркнуть важную задачу и потенциал камень преткновения для проведения теоретической редукции.
Шаффнер (1969) модифицировал модель Нагеля по включение идеи о том, что то, что на самом деле выводит теория восстановления (и, следовательно, объясняет) – это исправленная версия редуцированной теории, а не исходная теория.Он утверждал, что эта пересмотренная модель лучше зафиксированы сокращения по физическим наукам. Он потребовал своего исправленная модель может также использоваться, чтобы показать, как исправленная версия классическая генетика сводилась к новой физико-химической теории. наука называется молекулярной генетикой. Халл (1974) возражает против этого классического генетика не уменьшалась, по крайней мере, в соответствии с моделью сокращение, применяемое Шаффнером. Халл утверждал, что генетика не служат примером редукции Нагеля, потому что фундаментальные термины классической генетика не могла быть соответствующим образом связана с выражениями, выраженными в термины ДНК.
3.2 Антиредукционистский консенсус в отношении генетики
Большинство философов, пишущих о генетике и редукционизме, утверждали, что что молекулярная генетика не уменьшила и не уменьшит классическую генетику (например, см. Wimsatt 1976a, Darden and Maull 1977, Kitcher 1984, Розенберг 1985 и 1994, Дюпре 1993 и Буриан 1996). Два возражения против редукционистского тезиса Шаффнера были наиболее убедительно: возражение о несвязанности и кровавые подробности возражение. Возражение о несоединимости утверждает, что терминология классической генетики не может быть переопределена на молекулярном уровне. уровень с точки зрения ДНК.Это возражение фактически утверждает, что Второе формальное требование Нагеля – возможность подключения требование, не может быть выполнено. кровавые подробности возражение утверждает, что молекулярная генетика не может и не будет объяснить классическую генетику или лучше объяснить явления, которые уже объяснено классической теорией. Это возражение касается Первое формальное требование Нагеля, выводимость требование. Но возражение против кровавых деталей идет философски. глубже, потому что это подразумевает, что даже если объяснительные принципы классическая генетика может быть выведена из объяснительных принципов молекулярная генетика, выводы не будут объяснительными.
Самая строгая формулировка возражения о несоединимости может быть найдено в ранних работах Розенберга, который однажды утверждал, что представляет собой непреодолимый концептуальный разрыв между классическим и молекулярным теории генетики (1985, 1994). В поддержку этого утверждения, Розенберг утверждал, что отношения между генной концепцией классического генетика и концепции молекулярной генетики безнадежно сложные отношения «многие-многие», которые навсегда сорвать любую попытку систематически связать две теории.Розенберг начал свой анализ с того, что в классической генетика, гены идентифицируются по их фенотипическим эффектам. Классические генетики определили ген пурпурного цвета глаз, Например, путем проведения тщательно спланированных экспериментов по разведению и отслеживая распределение фенотипов цвета глаз в последовательных поколения лабораторной популяции. Причина классической генетики никогда не будет сведена к науке молекулярного уровня, согласно Розенберг (1985), заключается в том, что нет управляемой связи между концепция менделевского фенотипа и молекулярного гена:
Путь к образованию пигмента красных глаз начинается во многих различных молекулярных генов и проходит через несколько альтернативных разветвленных пути.… Путь от [молекулярных] генов также содержит избыточные, неоднозначные и взаимозависимые пути. Если мы дадим биохимическая характеристика гена красного цвета глаз либо апеллировать к частям своего пути синтеза или апеллировать к сегментов ДНК, с которых он начинается, наше молекулярное описание этого ген будет слишком сложным, чтобы дать какое-либо практическое объяснение результат. (Розенберг, 1985, с. 101)
Розенберг пришел к выводу, что, поскольку связь между молекулярными генами и менделевский фенотип чрезвычайно сложен, связь между любой молекулярной концепцией и концепцией менделевского гена также должны быть чрезвычайно сложным, тем самым блокируя любые систематические, редуктивные объяснение классической генетики на молекулярном уровне теория.
Кровавые подробности возражения можно проследить до писаний Патнэм (1965) и Фодор (1968), которые выступали против редукционизма ментальный на основании того, что психологические функции многократно реализованный. Это возражение против редукционизма было дальнейшим разработан в контексте генетики, наиболее тщательно Китчером (например, см. Kitcher 1984, 1999, 2001). Вслед за Халлом Китчер предполагает что классическая генетика – это генетика передачи. Классический теория, согласно Китчеру, объясняет передачу фенотипических черты характера, а не развитие фенотипических черт у индивидуума организмы.А явления передачи, по мнению Китчера, лучше всего объяснил на уровне цитологии: «Распределение генов гаметам нужно объяснять, а не репетировать кровавые детали перетасовка молекул, но благодаря наблюдению, что хромосомы выровнены попарно незадолго до мейотического деления, и что одна хромосома из каждой согласованной пары передается каждому гамета. ” (Китчер 1984, с. 370). Китчер предполагает, что спаривание и разделение хромосом относятся к естественному виду пары процессы разделения, которые являются гетерогенными от молекулярного перспектива, потому что разные силы несут ответственность за объединение и разъединение различных парных сущностей.В он утверждает, что разделение парных сущностей «может произойти из-за действие электромагнитных сил или даже ядерных сил; но это легко придумать примеры, в которых разделение осуществляется действие силы тяжести ». (Китчер 1984, с. 350)
Образ генетики, исходящий от антиредукционистов литература – это двухуровневая наука, состоящая из двух незаметных теоретические дискурсы, основанные на принципах о сущностях в цитологический уровень (например, хромосомы) и другой, основанный на принципы о сущностях на молекулярном уровне (например, нуклеотид последовательности в ДНК).Антиредукционисты верят в некоторые явления, включая передачу генов, лучше всего объясняются теорией обоснованы на цитологическом уровне и другие явления, в том числе экспрессии генов, лучше всего объясняются теорией, основанной на молекулярный уровень. Хотя Китчер утверждает, что классическая генетика дает лучшее объяснение в объективном смысле, некоторые антиредукционисты (например, Rosenberg 1985, 1994) считают, что Препятствия на пути сокращения являются чисто практическими . Розенберг (1985, г. 1994) обратился к концепции супервентности, чтобы доказать, что в принцип , молекулярная генетика обеспечит лучшее объяснения.Но он утверждал, что на практике , классический генетика дает «лучшее» объяснение передачи явления в том смысле, что это лучшее объяснение, доступное для существа с нашими когнитивными ограничениями. Однако впоследствии Розенберг изменил свою позицию по этому вопросу, в основном, на том основании, что что технологические достижения в хранении и обработке информации “может существенно повысить нашу способность понимать макромолекулярные процессы и их комбинации »(Розенберг 2006, стр.14).
Несмотря на философски значимые различия во взглядах на конечная основа несводимости классической генетики, образ биологического знания, возникающий из писания антиредукционистов похожи. Биологический мир состоит из разных областей явлений, и каждая область лучше объясняется на определенном уровне теоретического дискурса. Следовательно идеальная структура для биологии сродни слоеному пирогу с ярусами теории, каждая из которых дает наилучшее из возможных объяснений своих область явлений.Биологические науки, такие как классическая генетика которые сформулированы в терминах более высоких уровней организации, должны упорствовать, в безопасности от редуктивного понимания молекулярной науки, потому что их центральные теории (или модели рассуждений) объясняют области явления, которые лучше всего объясняются на уровнях выше, чем молекулярный уровень.
3.3 Вызовы антиредукционистскому консенсусу
Консенсус антиредукционистов не остался без ответа (см. 1989, 1992 и 1998, Schaffner 1993 и Waters 1990 и 2000).По мнению критиков, основные возражения в поддержку консенсуса ошибаются. Возражение о несоединимости основывается на предположении что классическая генетика рассматривала отношения между генами и фенотипические черты должны быть простыми отношениями один-к-одному. Но классический генетики знали лучше. Подумайте, что Стертевант, один из звездные студенты и сотрудники, должны были сказать о генах и глазах цвет:
Разница между нормальными красными глазами и бесцветными (белыми) у Drosophila происходит из-за различия в одном гене.Еще красный очень сложный цвет, требующий взаимодействия как минимум пяти (и, вероятно, очень многих других) разные гены его производство. И эти гены совершенно независимы, каждая хромосома несущие некоторые из них. Более того, цвет глаз косвенно зависит от большое количество других генов, таких как те, на которых жизнь летать зависит. Тогда мы ни в коем случае не можем идентифицировать данный ген с красный цвет глаз, хотя есть единственный ген отличая его от бесцветного глаза .Так это для всех персонажи … (выделено мной, цитата из Карлсона 1988, стр. 69)
Эта цитата предполагает, что связь между геном и цвет глаз в классической генетике проявлял ту же сложность, что и Розенберга обсуждались на молекулярном уровне (сравните эту цитату с отрывок из Розенберга 1985, цитируемый в разделе 3.2). В соответствии с эта критика возражения о несоединимости, это не так что отношения генотип-фенотип кажутся простыми и единообразными на уровень классической генетики и сложный и разрозненный на уровне молекулярный уровень.Ситуация выглядит одинаково сложной на обоих уровнях. анализа (Waters 1990).
Тем не менее классическая генетика находит простой способ объяснить явления передачи, обращаясь к принципу различия, в соответствии с какими конкретными различиями в конкретных генах вызывают особые различий фенотипических признаков у конкретные контексты (см. раздел 2.1). Стертевант намекает на это принцип в первом предложении цитаты выше и снова в подчеркнутый пункт.Возникает вопрос, могут ли эти отношения быть захваченным на молекулярном уровне? И ответ – да. В различия, используемые классическими генетиками для объяснения наследования паттерны обычно идентифицируются на молекулярном уровне с помощью современные генетики.
Согласно этой критике, возражение против кровавых деталей также не оправдывается. Это возражение утверждает, что биологи не могут улучшить классические объяснения явлений передачи, цитируя молекулярные детали. Цитологический уровень якобы обеспечивает наилучшее уровень объяснения, потому что объяснения на этом уровне единообразно учитывать широкий спектр случаев, которые выглядели бы неоднородными с точки зрения молекулярная перспектива.Рассмотрим формулировку Китчера этого возражение. Китчер считает, что объяснить – значит объединить (1989). Это следует, что лучшее объяснение класса явлений – это объяснение, которое описывает класс единообразно. Китчер утверждает, что мейоз является примером такого рода ситуации. Единообразие процессы разделения пар очевидны на цитологическом уровне, но теряется в кровавых деталях на молекулярном уровне, где процесс «Может произойти из-за действия электромагнитных сил или даже ядерных сил… »(Китчер, 1984, с.350). Но неясно, что мог иметь в виду Китчер. Молекулярный механизмы, лежащие в основе спаривания и разделения хромосом: удивительно однородны по животным, от дрожжей до людей; Это Дело не в том, что некоторые из них связаны с электромагнитными силами, а другие задействовать ядерные силы. Заявление Китчера о том, что «это легко подумать о примерах, в которых разделение осуществляется действием гравитации »не имеет оснований в том, что молекулярные биологи узнал о спаривании и разделении хромосом.
Мейоз – бесперспективный кандидат для иллюстрации идеи того, что кажется единообразным на уровне классической генетики оказывается неоднородны на молекулярном уровне. Но эту идею иллюстрирует другие генетические явления. Рассмотрим феномен генетического доминирования. В классической генетике рассматриваются все примеры полного доминирования. одинаково для целей объяснения явлений передачи. Но современная генетика показывает, что существует несколько очень разных механизмы, лежащие в основе различных примеров доминирования.В соответствии с Унификационистская теория научного объяснение, классический подход к доминированию дает объективное лучшая основа для объяснения явлений передачи, потому что она обеспечивает более унифицированная организация явлений. Но это означало бы, что поверхностные объяснения классической генетики объективно предпочтительнее более глубоких объяснений, предоставляемых молекулярной теорией (Уотерс 1990).
3.4 Выход за рамки теоретической редукции и слоеного антиредукционизма
Хотя концепция теоретической редукции Нагеля отмечает общий отправной точкой для дискуссий о очевидном сокращении классической генетики, большая часть литературы по редукции направлена на стремясь лучше понять природу редукции, ища заменить концепцию Нагеля на более информативную.Это правда антиредукционистов, которые стремятся выяснить, почему молекулярная генетика не может уменьшить классическую генетику, а также тех, кто был более сочувствует редукционизму. Следовательно, существует два уровня дискурса. в литературе, исследующей вопрос о том, является ли молекулярная генетика сокращает классическую генетику. Один уровень касается того, что происходит в науке генетики. Другой касается более абстрактных вопросов. о природе (гносеологической) редукции.
Абстрактный уровень дискурса начался с идеи Шаффнера о том, что сокращена не исходная теория, а исправленная версия исходной теории.Wimsatt (1976a) предлагает более амбициозный модификация. Он отвергает предположение, что научные теории наборы утверждений, подобных закону, и что объяснения являются аргументами в Объясняемые явления вытекают из законов. Вместо того опираясь на эти предположения, Вимсатт использует версию Сэлмона о объяснение (Salmon 1971), чтобы проверить утверждения о том, что молекулярная генетика предложил редуктивные объяснения. Китчер (1984) также отвергает учет теоретизирования, лежащего в основе концепции редукции Нагеля.Он строит новую концепцию редуктивного объяснения, основанную на его собственном представление о том, что эффективно составляет научную теорию, и его унификационистский взгляд на научное объяснение (1989). Так же, Саркар (1998) отвергает теорию и объяснение. предполагается в концепции редукции Нагеля. Фактически, он явно избегает полагаться на какой-либо конкретный отчет научных теорий или теоретическое объяснение. Вместо этого он предполагает, что редуктивная объяснения – это объяснения без указания, что за объяснение есть, а затем пытается определить особенности, которые устанавливают редуктивные объяснения помимо других объяснений.
Вимсатт, Китчер и Саркар стремятся заменить концепцию Нагеля о редукция с концепцией, которая не предполагает, что научный объяснение предполагает отнесение к универсальным законам. Вебер (2005), однако стремится заменить концепцию Нагеля концепцией, в которой сохраняется идея о том, что редуктивное объяснение включает в себя отнесение к законам сокращение науки. Вебер отвергает идею о том, что редукционизм в Биология включает объяснение биологических законов более высокого уровня. Он утверждает что, за некоторыми редкими исключениями, в биологических науках нет законы.Он утверждает, что редукционизм в биологии предполагает объяснение биологические явления непосредственно в терминах физических законов. Следовательно, он отвергает “слоеную” концепцию редукции, заложенную в концепции Нагеля. учетная запись.
Литература по восстановлению и молекулярной генетике оказала влияние на мысли философов о редукции в других науках. Например, Концепция восстановления Китчера, которую он использует, чтобы объяснить, почему молекулярная генетика не может уменьшить классическую генетику, впоследствии была использовала Хардкасл (1992) в ее исследовании отношений между психологией и нейробиологией.С другой стороны, Sober развивает и расширяет критику возражения Китчера о кровавых деталях (раздел 3.3), пересматривая аргументы Патнэма (1967, 1975) и Фодора. (1968, 1975) о множественной реализуемости.
Собер (1999) утверждает, что науки более высокого уровня могут описывать закономерности. невидимы на более низком уровне и, следовательно, могут предлагать более общие объяснения. Но он настаивает на том, что описание не следует путать. с объяснением. Он утверждает, что хотя физика, возможно, не сможет в описать все шаблоны, тем не менее, он может объясните любое единичное явление, которое наука более высокого уровня могу объяснить.Науки более высокого уровня могут дать более «общие» объяснения, но физика дает «более глубокие». Он предлагает, чтобы какое объяснение лучше – в глазах смотрящего. Он заключает, что
… Редукционисты, возможно, захотят сдержаться в этом вопросе о лучшем и того хуже, и попытайтесь построить на простом предположении, что физика в принцип может объяснить любой единичный случай, когда высокоуровневый наука может объяснить. … Для редукционистов интересная особенность физических объяснений социальных, психологических, и биологических явлений заключается в том, что они используют те же основные теоретические машины, которые используются для объяснения несоциальных явлений, непсихологические и небиологические (Sober 1999, стр.560-1).
Обсуждение замкнулось. Множественная реализуемость аргумент, критикуемый Собером, был основан на абстрактных соображения в контексте философии разума. Философы биология обратилась к этой литературе, чтобы построить кровавые детали возражение против идеи, что молекулярная генетика сокращает классическая генетика. Другие философы утверждали, что это возражение действительно не выдерживают тщательного анализа конкретной ситуации в генетика. Собер извлек уроки из дискуссии о генетики, чтобы критиковать исходный аргумент, препятствующий реализации, и привлекать общие выводы о редукционизме.
В работах Вимсатта о редукции (1976a, 1976b и 1979) подчеркивается плодотворность попытки добиться сокращения, даже когда снижение не достигается. Он утверждает, например, что попытки раскрыть молекулярный состав сущностей, идентифицированных на более высоких уровнях уровни часто бывают плодотворными, даже когда идентичность между уровнями не может быть найденным. Кроме того, Wimsatt отмечает, что «затраты» разработки редуктивных объяснений многих деталей уже объясненные на более высоком уровне имеют отношение к вопросу о том, почему не является полноценной заменой объяснений более высокого уровня на нижнего уровня.Возможно, тот факт, что молекулярная генетика не замененная классическая генетика может быть объяснена на основе высоких издержки, а не отсутствие эпистемических достоинств.
Хотя Шаффнер по-прежнему утверждает, что молекулярная генетика может принцип сокращения классической генетики, он признал, что попытки выполнение сокращения будет «периферийным» по отношению к развитие молекулярной генетики. Можно было бы ответить примерно так: Халл (1977), что успех молекулярной генетики кажется редуктивный в некотором важном смысле.Следовательно, неспособность осветить этот успех с точки зрения редукции обнаруживает концептуальную несостоятельность. Что можно утверждать, что тезис о периферийности Шаффнера указывает что его концепция редукции не является эпистемически релевантной потому что он не может пролить свет на плодотворность редуктивного исследования в молекулярная генетика.
Фактически, общий недостаток дебатов о сокращении классическая генетика состоит в том, что она касается лишь фрагмента научных рассуждения. Он основан почти исключительно на анализе объяснительные или теоретические рассуждения и в значительной степени игнорируют следственные рассуждения.Философская литература о предполагаемом сокращении классическая генетика фокусируется на том, как генетики объясняют или пытаются объяснять явления, а не то, как они манипулируют или исследуют явлений. Это справедливо даже для Вимсатта (1976a). учет эвристик, которые используют эвристику для объяснения.
Вэнс (1996) предлагает более тщательный сдвиг внимания с теории на следственная практика. Он утверждает, что есть только один современник наука о генетике и описывает, как следственные методы классическая генетика является неотъемлемой частью методологии того, что называется молекулярной генетики .Он заключает, что редукционизм терпит неудачу, потому что современная генетика все еще зависит от методов классическая генетика, включающая селекционные эксперименты. Фотография Вэнса генетика убедительна. Лабораторные методы классической генетики действительно сохраняются, даже если они значительно расширяются, дополняются и часто заменяется методами прямого вмешательства в ДНК. Но Картина Вэнса не соответствует антиредукционистскому образу двухуровневая наука и утверждение, что классическая генетика оставаться в стороне от редуктивного понимания молекулярной биологии.
Иная картина возникает, если рассматривать генетику как следственный наука, предполагающая взаимодействие методологических и объяснительных рассуждения (Waters 2004a). Этот образ не двухуровневой науки, один (классическая генетика), направленный на исследование и объяснение явления передачи и другое (молекулярная генетика), направленное на исследование и объяснение феноменов развития. Вместо этого там одна наука, которая сохраняет большую часть исследовательских и объяснительных рассуждения классической генетики путем переосмысления ее теоретических основы в молекулярных терминах и путем переоснащения его основных исследовательских подход путем интеграции методологий классической генетики с физически обоснованные методы биохимии и новые методы, основанные на рекомбинантные ДНК и технологии интерференции РНК.
Распространенное утверждение в философской литературе о молекулярных генетика состоит в том, что гены невозможно представить на молекулярном уровне. Из Конечно, философы не отрицают, что биологи используют термин , ген , но многие философы считают, что ген – пустышка. термин, заменитель множества различных понятий. Разные ответы генного скептицизма иллюстрируют множество философских целей и подходы. Один из вариантов ответа – внимательно проанализировать объяснения. связаны с экспериментальной практикой (а не с широкими обобщениями фундаментальной теории), чтобы определить, существуют ли однородные шаблоны рассуждений о генах, которые могут (а) быть кодифицированы в четкие концепции и / или (б) используются для установления ссылки на срок.Другой вид ответа – предложить новые концепции генов, которые лучше послужит поставленным целям практикующих биологов. Треть своего рода ответ заключается в проведении анализа опроса, а не в проведении традиционные методы философского анализа. Четвертый вид ответ состоит в том, чтобы принять (предположительно) необходимую неопределенность гена концепции и изучить, почему использование термина ген так полезный.
4.1 Генный скептицизм
Генные скептики утверждают, что путь непоследователен. Ген используется на молекулярном уровне, и что этот термин не обозначать натуральный вид; скорее, якобы используется ген чтобы выделить много разных единиц в ДНК.ДНК состоит из «Кодирующие» области, которые транскрибируются в РНК, разные видов регуляторных областей, а у высших организмов ряд регионы, функции которых менее ясны и, возможно, в случаях не существует. Скептицизм в отношении генов частично основан на идее, что этот термин иногда применяется только к частям области кодирования, иногда для всей области кодирования, иногда для частей кодирования регион и регионы, которые регулируют этот кодирующий регион, а иногда ко всей кодирующей области и регуляторным регионам, влияющим или потенциально влияя на транскрипцию кодирующей области.Скептики (например, Burian 1986, Portin 1993 и Kitcher 1992) заключил, как лаконично выразился Китчер: «ген – это то, что компетентный биолог предпочитает называть ген »(Kitcher 1992, п. 131).
Учебники по биологии содержат определения гена, и это поучительно рассмотреть один, чтобы показать, что концептуальные ситуация действительно тревожная. Самый распространенный современный определение состоит в том, что ген – это фундаментальная единица, которая кодирует полипептид. Одна проблема с этим определением заключается в том, что оно исключает многие сегменты, которые обычно называют генами.Некоторые Сегменты ДНК кодируют функциональные молекулы РНК, которые никогда не переводится в полипептиды. Такие молекулы РНК включают РНК переноса, рибосомная РНК и молекулы РНК, которые играют регуляторную и каталитическую роли. Следовательно, это определение слишком узкое.
Еще одна проблема с этим общим определением заключается в том, что оно основано на чрезмерно упрощенное описание экспрессии ДНК. Согласно этому Проще говоря, ген – это последовательность нуклеотидов в ДНК, которая транскрибируется в последовательность нуклеотидов, составляющих информационную РНК молекула, которая, в свою очередь, транслируется в последовательность аминокислот, которая образует полипептид.(Биологи говорят так, будто гены «производят молекулы полипептида »или« предоставляют информацию для полипептида ».) Реальная ситуация с экспрессией ДНК, однако часто бывает гораздо сложнее. Например, в растениях и животных, многие молекулы мРНК обрабатываются, прежде чем они будут переведены в полипептиды. В этих случаях части молекулы РНК, называется интронов , отрезаны и оставшиеся сегменты, называется экзонов , сплайсируются до молекулы РНК покидает клеточное ядро.Иногда биологи называют всю ДНК область, которая соответствует как интронам, так и экзонам, ген. В других случаях они называют только части сегмента ДНК соответствующие экзонам гена. (Это означает, что некоторая ДНК сегменты, которые генетики называют генами, не являются непрерывными сегментами ДНК; они представляют собой наборы прерывистых экзонов. Генетики звонят эти расщепляют гены .) Дальнейшие осложнения возникают из-за того, что сплайсинг экзонов в некоторых случаях выполняется по-разному в разные типы тканей и на разных стадиях развития.(Этот означает, что имеется перекрывающихся генов .) Проблема с общее определение, что гены – это сегменты ДНК, которые «кодируют полипептиды »заключается в том, что понятие« кодирование полипептид »неоднозначен, когда речь идет о реальных осложнениях. экспрессии ДНК. Генные скептики утверждают, что это безнадежно неоднозначно (Burian 1986, Fogle 1990 и 2000, Kitcher 1992 и Portin 1993).
Ясно, что это определение, которое является наиболее распространенным и заметным определение из учебника слишком узкое, чтобы его можно было применять к диапазону сегменты, которые генетики обычно называют генами и слишком неоднозначны, чтобы обеспечить единое точное разделение ДНК на отдельные гены.Учебники включают множество определений гена. По факту, философов часто разочаровывала тенденция биологов определить и использовать термин ген в ряде противоречивых способов в одном и том же учебнике. После рассмотрения альтернативы определения к философскому исследованию, генные скептики пришли к выводу что проблема не просто в отсутствии аналитической строгости. В проблема в том, что гена просто не существует. молекулярный уровень. То есть нет единого, единообразного и однозначного способ разделить молекулу ДНК на разные гены.Генные скептики часто утверждали, что биологи должны излагать свою науку в терминах Сегменты ДНК, такие как экзон, интрон, промоторная область и т. Д., И полностью отказаться от термина ген (наиболее убедительно аргументировано Fogle 2000).
4.2 Анализ концепций на практике,
классический ген и молекулярный ген концепцииПротив генного скептицизма утверждалось, что биологи последовательный, точный и единообразный способ представления генов на молекулярный уровень.Анализ, лежащий в основе этого аргумент начинается с различения двух разных способов современные генетики думают о генах. Классические генетики часто воспринимаются гены как функциональные единицы в хромосомах, различия, которые вызывают различия в фенотипах. Сегодня в контексты, в которых гены идентифицируются по наблюдаемым фенотипическим признакам. различия, генетики до сих пор представляют себе гены в этом классическом образом, как функциональные единицы в ДНК, различия которых вызывают наблюдаются различия в фенотипах.Такой способ представления генов назвал классическую концепцию гена (Waters 1994). Но современные генетики также иначе думают о генах, ссылаясь на концепцию молекулярного уровня. Молекулярный ген Концепция основана на идее, что гены – это единицы ДНК, которые функция для определения линейных последовательностей в молекулах, синтезированных с помощью Экспрессия ДНК. Согласно этому анализу, обе концепции работают. у современных генетиков. Moss 2003 также различает два современные концепции генов, которые он называет «гены-P (преформистский) »и« гены-D (онтогенетические) ».Он утверждает, что смешение этих понятий приводит к ошибочному мышлению в генетике.
В классическом представлении о генах много путаницы. из-за того, что генетики иногда говорили так, как будто классически задуманные гены – для общего фенотипа признаки (фенотипы) или как будто отдельные гены производят фенотипы. Этот разговор был очень вводящим в заблуждение со стороны классических генетиков и продолжает вводить в заблуждение в контексте современная генетика.Производство грубого фенотипического признака, например, фиолетовый цвет глаз, включает в себя все виды генетических и внегенетические факторы, включая различные клеточные ферменты и структуры, устройства тканей и факторы окружающей среды. В кроме того, неясно, каков общий фенотипический уровень, если таковой имеется. функции можно отнести к отдельным генам. Например, нет сегодня яснее, чем во времена Моргана, что функция фиолетовый ген, обсуждаемый в разделе 2.1, вносит свой вклад в изготовление цвета глаз.Мутации в этом гене влияют на ряд признаки общего фенотипического уровня. Законное объяснительное рассуждение обращение к классической концепции гена не зависит ни от какого багажа относительно того, для чего предназначены гены или какую функцию ген может выполнять в разработка. От чего зависит объяснительное рассуждение, так это от принцип различия, то есть принцип, согласно которому некоторая разница в ген вызывает определенные фенотипические различия в конкретных генетических и экологический контекст (раздел 2.1). Множество объяснений, основанных на генах в современной биологии лучше всего понимаются с точки зрения классических генная концепция и принцип различия.
Возможно, причина, по которой генные скептики упустили из виду концепцию молекулярных генов в том, что они искали неправильную концепцию. Концепция не является чисто физико-химической концепцией и не дает однократное разделение ДНК на отдельные гены. Вместо этого это функциональная концепция, которая обеспечивает единообразное представление о генах которые можно применять для выделения разных сегментов ДНК в разных следственный или объяснительный контекст. Базовый молекулярная концепция, согласно этому анализу, – это концепция ген линейной последовательности в продукте экспрессии ДНК:
Ген g для линейной последовательности l в продукте p синтезируется в клеточном контексте c является потенциально реплицирующаяся нуклеотидная последовательность, n , обычно содержащаяся в ДНК, что определяет линейную последовательность l в продукте p на некоторой стадии экспрессии ДНК (Waters 2000)
Понятие молекулярного гена можно представить в виде 4-кортежа: < n , l , p , c >.Этот анализ показывает, как генетики могут последовательно включать интроны как часть ген в одном эпистемическом контексте, а не в другом. Если контекст включает идентификацию гена первичной, предварительно обработанной молекулы РНК, тогда ген включает как интроны, так и экзоны. Если контекст включает идентификацию гена полученного полипептида, тогда ген включает только экзоны. Следовательно, в случае ДНК выражение, которое в конечном итоге приводит к синтезу данного полипептид, генетики могут говорить так, будто «ген» включены интрон (в этом случае они будут относиться к гену для первичной, предварительно обработанной РНК ), но при этом говорить так, как если бы «Ген» исключил интроны (в этом случае они относится к гену зрелой РНК или полипептид ).Применение концепции молекулярного гена не является двусмысленный; на самом деле, это замечательно точно, если указать значения переменных в выражении «ген для линейных последовательность l в продукте p синтезируется в клеточном контекст c ».
Генные скептики предположили, что в генах отсутствует согласованность. говорить, потому что биологи часто говорят так, будто гены кодируют полипептиды, но затем развернитесь и поговорите о генах молекул РНК, которые не транслируется в полипептиды (включая гены РНК [тРНК], рибосомная РНК [рРНК] и интерференционная РНК [iRNA]).Эта учетная запись показывает что зачатие генов рРНК включает в себя ту же идею, что и зачатие генов полипептидов. В обоих случаях ген является сегментом ДНК, расщепленная или нет, которая определяет линейную последовательность в молекуле представляет интерес.
Преимущество этого анализа состоит в том, что он подчеркивает ограничения геноцентричных объяснений при уточнении характерных причинных роль генов в синтезе РНК и полипептидов: гены определить линейные последовательности первичных транскриптов РНК и часто играют особую роль, хотя и не исключительную, в определении последовательность аминокислот в полипептидах.
4.3 Практический анализ ссылки, как молекулярная обработка изменила ссылку
Вебер (2005) исследует эволюцию концепции гена, отслеживая изменения в упоминании термина ген в истории генетики. Ссылка или продление срока – набор объектов, на которые он ссылается. Вебер придерживается смешанной теории рефенс. Согласно смешанным теориям, термин определили, как соответствующее языковое сообщество причинно взаимодействует с потенциальными референтами, а также как они описывают потенциальные референты.Эта теория заставляет Вебера уделять пристальное внимание, а не только как генетики теоретизировали о генах или использовали эту концепцию для объяснения явлений, но также и то, как они проводили свою лабораторию расследования. Следуя Китчеру (1978, 1982), он исследует способы какие способы обращения менялись с течением времени.
Вебер выделяет шесть различных концепций генов, начиная с теории Дарвина. пангене (1868 г.) и заканчивая современной концепцией молекулярная генетика. Он выделяет современные молекулярные от классической (или «неоклассической») концепции основа того, как генетики описали их функциональную роль (РНК / белок кодирование по сравнению с общей функциональной единицей), их материальная основа (РНК / ДНК по сравнению с хромосомой), и их структура (прерывистая линейная – с интронов и экзонов по сравнению с непрерывными линейными), а также на основе критерии, используемые экспериментаторами для идентификации генов (по продукту гена по сравнению с тестом на комплементарность).
Вебер исследует, как расследование нескольких частных Drosophila генов изменилось как наука генетика развитый. Его исследование показывает, что методы молекулярной генетики предоставили новые способы идентификации генов, которые были впервые идентифицированы классические техники. Упоминание термина изменилось не просто как в результате теоретических разработок, но также и в результате внедрение новых методов идентификации генов. Он заключает, что в отличие от концепций физической науки, которые были проанализированы философов, концепция гена имеет «несенциалистский характер, который позволяет биологам устанавливать различные природные классификации, в зависимости от доступных методов расследования, а также от теоретические интересы »(Weber 2005, p.228). Вебер называет эту функцию «Плавающие ссылки».
4.4 Предложение по новой концепции, технологии молекулярного гена
концепцииNeumann-Held (2001) предлагает новый взгляд на гены в контекст генетики развития. Она говорит, что в этом контексте интерес к генам в основном сосредоточен на регулируемой экспрессии полипептиды. Она отмечает, что определения гена из учебников часто признаем этот интерес и цитируем следующее определение из научный учебник:
Комбинация сегментов ДНК, которые вместе составляют экспрессируемый единица, выражение, ведущее к образованию одного или нескольких конкретных функциональные генные продукты, которые могут приводить либо к молекулам РНК, либо к полипептиды.Сегменты гена включают (1) транскрибируемую единицу … И любые регуляторные сегменты, включенные в транскрипцию единица, и (2) регуляторные последовательности, фланкирующие транскрипцию единицы и требуются для конкретного выражения. (Сингер и Берг 1991, п. 41).
Это определение подчеркивает, что регуляторные последовательности, а также кодирующие области требуются для «конкретных выражение.” Лишь небольшая часть кодирующих последовательностей транскрибируются в данной ячейке в определенное время, и независимо от того, конкретная последовательность транскрибируется, частично зависит от регуляторных регионы, внешние по отношению к кодирующей области.
Нойман-Хельд указывает, что если цель состоит в том, чтобы указать, что необходим для регулируемого синтеза полипептидов, то необходимо включать даже больше, чем то, что находится в ДНК. Это следует из тот факт, что такие процессы, как дифференциальный сплайсинг (и редактирование РНК процессы, такие как метилирование, которые я не обсуждал в этом article) вовлекают сущности за пределами ДНК, такие как агенты сплайсинга. Она предполагает, что это уместно, по крайней мере, в контексте генетика развития, чтобы переосмыслить гены как процессы.Она предлагает следующая концепция молекулярного гена процесса .
«Ген» – это процесс (т. Е. Ход событий), который связывает вместе ДНК и все другие соответствующие объекты, не относящиеся к ДНК, в производство определенного полипептида. Термин ген в этом смысл обозначает процессы, которые определяются (1) конкретным взаимодействия между определенными сегментами ДНК и определенными не ДНК локализованные объекты, (2) конкретные механизмы обработки результирующих мРНК во взаимодействии с дополнительными объектами, не локализованными в ДНК.(Нойман-Хельд 2001, стр.74).
Нойман-Хельд утверждает, что эта концепция дает наиболее четкую основу. для понимания того, как последовательности ДНК «используются в процессах производства полипептида ». Она указывает, что концепция молекулярного гена процесса позволяет включать кодирование последовательности в ДНК, регуляторные последовательности в ДНК, а также объекты, не расположены в ДНК, и все они причинно участвуют в производстве полипептидов. Концепция Неймана-Хельда исключает транскрипцию процессы и кодирующие области ДНК, которые приводят к функциональной РНК молекулы, которые не транслируются в полипептиды.Следовательно, согласно по ее мнению, не существует процессных молекулярных генов для тРНК. (транспортная РНК), рРНК (рибосомная РНК) или мяРНК (малые ядерные РНК). Эта особенность определения Неймана-Хельда не соответствует определение из учебника, которое она цитирует, чтобы мотивировать свою точку зрения (представлено выше). Кроме того, исключение этих кодирующих областей не трек с недавними открытиями о важных функциях, выполняемых некодирующие молекулы РНК, такие как мяРНК. Ее определение легко могло быть должны быть пересмотрены с учетом этих регионов и процессов.В любом слючае, Нойман-Хельд считает, что использование этой концепции в генетике развития, а не концепции генов, ориентированных на ДНК, поможет избежать мнения, что «Гены – самые важные объясняющие факторы в биологии. из-за их уникальной причинной силы »(Neumann-Held 2001, п. 80).
4.5 Использование подхода на основе опросов для определения концепций генов
Стотц и Гриффитс (2004) считают, что разнообразие концепций генов используется во всех биологических науках и требует более систематического и явно эмпирический подход.Они указывают на этого человека философы не могут понять всех тонкостей различных контексты в широком спектре биологических наук, в которых ген используются концепции. Они приступили к амбициозному проекту, чтобы опросить практикующих ученых в попытке определить, как Ученые на самом деле задумываются о генах. Их интерес расширяется далеко за пределами понимания «молекулярной генетики». Они надеются узнать о концепциях, используемых во многих различных областях и контекстов биологии, обнаруживая различия в способах, которыми биологи разные области (и биологи разных возрастных групп, полов и т. д.) отвечать на сложные анкеты.
Первоначальной мотивацией проекта Стотца и Гриффита было испытание философское рассмотрение концепции гена. Как спросил Гриффитс, если их исследование, основанное на опросе, показало, что на самом деле ученые не подумайте о генах так, как это изложено в философском изложении, затем какую ценность может иметь учетная запись? Однако есть количество серьезных практических трудностей с использованием анкеты чтобы узнать, как думает человек, особенно если он думает предполагает использование нескольких концепций и / или иногда или в некоторой степени запутанный (Waters 2004b).Также сложно определить подходящие и репрезентативные выборки ученых. Гриффитс и Стотц знают о эти трудности и усовершенствовали свой проект через последовательные опросы.
Даже если исследование Стотца и Гриффита поможет определить, как ученые в разных областях биологии действительно думают о генах в в разных контекстах, из этого не следует, что их выводы будут обеспечить соответствующий тест классического, молекулярного или технологического концепции молекулярных генов.Цель сторонников этих концепций состоит в том, чтобы переосмыслить знания современной генетики, заменив небрежное мышление, основанное на неясных концепциях, с более строгим мышлением с точки зрения точных понятий. Показывая, что фактические учёные мышление не согласуется с точным применением этих концепций не будет опровергать анализ, подтверждающий классический ген, или концепции молекулярных генов, и это не подорвет аргумент мотивируя предложение о новом процессе молекулярного гена концепция.
Хотя кажется, что результаты опроса не дадут соответствующий тест философского анализа концепций генов, они может предоставить, как утверждают Стотц и Гриффитс, важную информацию актуально для тех, кто проводит философские исследования концепций генов. Например, если такие опросы обнаруживают существенные различия в способах эволюционные биологи и генетики развития отвечают на вопросы о том, что считается геном, философы могли бы исследовать, контексты, в которых практикуют эти биологи, требуют различных генов концепции.Результаты опроса могут предоставить полезную эвристику для проведение концептуального анализа.
4.6 Прагматический ответ на генный скептицизм
Генные скептики, такие как Буриан, Портин и Фогл, утверждают, что термин Ген изжил себя. Они утверждают, что термин и слишком расплывчато, и слишком ограничительно. Они считают, что это слишком расплывчато, потому что он не обеспечивает однозначного анализа генома. Границы между генами перекрываются и якобы неоднозначны. Нет ясно, спорят они, включают ли гены интроны или исключают их, регулирующие регионы и т. д.Термин якобы тоже ограничительный, потому что он скрывает разнообразие молекулярных элементов играют разные роли в экспрессии и регуляции ДНК. В Кроме того, при любых попытках разрешить двусмысленность эти скептики поспорим, сделаю термин еще более ограничительным.
Отчет Келлера об истории генетики двадцатого века, кажется, усиливают генный скептицизм. Например, она утверждает, что вопрос о том, для чего нужны гены, становится все труднее отвечать (Келлер 2000).К концу двадцатого века, по ее словам, биологические находки выявили сложность развития динамика, из-за которой невозможно представить себе гены как отдельные возбудители в развитии. Келлер подчеркивает, что слова имеют силу и уделяет много внимания тому, как бессвязный разговор о генах повлияли на биологические исследования, укрепив предположение, что ген – это «основная объяснительная концепция биологической структуры. и функция »(Keller 2000, p. 9), допущение, с которым она категорически не согласен.Тем не менее Келлер не поддерживает точку зрения о генах. скептики, которые утверждают, что биология улучшится, если биологи перестали говорить о «генах» и ограничились термины, обозначающие молекулярные единицы, такие как нуклеотид, кодон, кодирование регион, промоторный регион и т. д. Келлер утверждает, что термин Ген продолжает иметь «очевидные и неоспоримые использует. ”
Согласно Келлеру, термин ген используется в том, что его расплывчатость, та самая черта, которая беспокоит философов, делает ее возможно, чтобы биологи были гибкими, чтобы общаться через дисциплинарные границы и мыслить по-новому:
Смысл экспериментального эффекта зависит от его отношения к другие эффекты и использование языка, слишком тесно связанного с конкретными экспериментальные практики в силу своей специфики эффективное общение в разных экспериментальных контекстах невозможно.(Келлер, 2000, с. 140).
Келлер указывает на вторую причину, по которой разговор о гене полезен. Термин , ген применяется к объектам, которые могут быть экспериментально манипулируют для получения определенных и воспроизводимых эффектов (хотя Келлер критикует концепции генов, неясно, какие сущности она думает, что этот термин относится). Она предполагает, что гены недолговечны. причины. Однако она отмечает, что это не означает, что гены долгосрочные причины или что гены являются основными причинными агентами разработка.Скорее, то, что это означает (а Келлер думает, что это важная причина, по которой разговор о генах будет продолжаться) в том, что гены могут быть используется в качестве ручек для управления биологическими процессами (см. также Воды 2000). По этим двум причинам, заключает Келлер, генный разговор будет и должен продолжать играть важную роль в биологический дискурс.
Наука под названием молекулярная генетика связана с фундаментальным теория, согласно которой гены и ДНК направляют всю основную жизнь процессов, предоставляя информацию, определяющую разработку и функционирование организмов.Говорят, что геном определяет «Программа развития», «генеральный план» или «Чертеж» для развития, в то время как другие элементы обеспечивают материалы (например, Bonner 1965, Jacob and Monod 1961, Mayr 1961, Мейнард Смит 2000, Розенберг 2006). Хотя идея, что хромосомы содержат «код-сценарий» для развития и функционирование организма было классно выражено Шредингером. (1944) до эпохи молекулярной генетики, сегодня часто выражены в явно молекулярных терминах. Информация о развитие и функции, которые передаются из поколения в поколение следующий, как утверждается, кодируется нуклеотидными последовательностями, содержащими гены и ДНК.Эта так называемая «генетическая информация» сначала «транскрибируется» в РНК, затем «Переведены» в белки, и, наконец, «Выражается» в развитии и функционировании организмы.
Концепция генетической информации занимает видное место в история молекулярной генетики, начиная с Уотсона и Крика. наблюдение, что, поскольку любая последовательность пар нуклеотидных оснований может соответствовать в структуру любой молекулы ДНК, «что в длинной молекуле возможно много разных перестановок, и поэтому кажется вероятно, что точная последовательность оснований – это код, который несет генетическую информацию.»(Уотсон и Крик, 1953). В качестве Как рассказывает Даунс (2005), генетики Джейкоб и Моно усилили использование информационного языка, как и те, кто пытался взломать “генетический код”. К началу 1960-х годов язык информация прочно закрепилась в области молекулярных генетика.
Философы в целом критиковали теорию о том, что гены и ДНК предоставить всю информацию и оспорить использование подметания такие метафоры, как «генеральный план» и «программа» которые предполагают, что гены и ДНК содержат все Информация.Критики заняли разные позиции. Самый похоже, принимают идею о том, что биологические системы или процессы содержат информации, но они отрицают идею о том, что ДНК играет исключительную роль в предоставление информации. Некоторые довольствуются утверждением, что при различных существующие теории информации, такие как причинные теории или стандартные телеосемантические теории, информация не ограничивается ДНК. Но другие утверждают, что понимание того, что делают гены, требует нового представление о биологической информации.Один из подходов – отступить к узкая концепция кодирования, специально направленная на прояснение смысла в котором ДНК предоставляет информацию для синтеза полипептидов, но не для черт более высокого уровня (например, Годфри-Смит 2000). Другой подход заключается в построении новой, широкой концепции биологических информации и используйте эту концепцию, чтобы показать, что информационная роль генов не исключительна (Яблонька 2002). Другой подход состоит в том, чтобы полностью отказаться от информационных разговоров и объяснить следственные и объяснительные рассуждения, связанные с генетикой и молекулярная биология с чисто причинной точки зрения.
5.1 Критика идеи о том, что гены и ДНК предоставляют информацию
Фундаментальная теория, гласящая, что роль ДНК заключается в обеспечении информация для развития подвергалась критике на многих основания. Келлер (2000) указывает, что эта идея двусмысленность. Предоставляет ли ДНК «программу» или “данные?” Другие утверждали, что «информация» для развития поступает из огромного количества «ресурсов», не только генетические ресурсы. Ояма (1985) предполагает, что это ошибка думать, что информация содержится в статических объектах, таких как ДНК.Она считает, что информация существует в жизненных циклах. Другой критика оспаривает применение определенных концепций или теории информации, включая приложения причинной и телеосемантические концепции.
Гриффитс (2001) различает два способа представить себе информации, причинной и преднамеренной, а затем утверждает, что либо зачатие, информация не ограничивается ДНК. Причинные теории информация, основанная на Dretske (1981), имеет отношение к Шеннону. математическая теория информации (1948).Дрецке отличает между исходной переменной и фоном или условиями канала. На Гриффитс (2001) прочтение теории Дрецке, исходной переменной, X, несет информацию о переменной Y , если значение X коррелирует со значением Y. Griffiths описывает причинную интерпретацию этой идеи следующим образом:
Существует канал между двумя системами, когда состояние одной систематически причинно связаны друг с другом, так что состояние отправителя можно обнаружить, наблюдая за состоянием получателя.В причинная информация – это просто состояние дел, с которым она надежно коррелирует на другом конце канала. Таким образом, дым несет информацию о фенотипах огня и болезней. информация о генах болезней. (Гриффитс, 2001, с. 397)
Чтобы уловить общепринятые представления о генетической информации в рамках этого теории, гены рассматриваются как исходные переменные, а среда – рассматривается как состояние канала. Отсюда следует, что гены несут информацию о фенотипах, потому что фенотипические значения надежно коррелируют с генотипические значения.Но, как отмечает Гриффитс, ничто не мешает рассматривая условия окружающей среды как исходные переменные, а гены как канал. В соответствии с этим применением причинной теории окружающая среда условия несут информацию о фенотипах. Гриффитс и другие пришли к выводу, что идея о том, что гены предоставляют информацию, в то время как другие причинные факторы просто предоставляют материал, который не может быть выдержан при причинные теории информации.
Гриффитс утверждает, что идея о том, что гены и ДНК обеспечивают все информация не лучше справляется с преднамеренными теориями информации.Интенциональные теории нацелены на то, чтобы уловить смысл «Семантическая информация», которую человеческие мысли и высказывания предположительно содержат (Годфри-Смит 1999). Версия умышленного Теория, которую предпочитают философы биологии, телеосемантична. Согласно для телеосемантических теорий сигнал представляет собой то, что он выбран для представления (в процессе эволюции). Согласно этой идее, можно сказать, что ДНК содержит информацию о развитии, потому что Влияние ДНК на развитие было выбрано в процессе эволюция.Но, как отмечают Гриффитс и Грей (1997), эта идея применяется к широкому кругу организаций, участвующих в разработке, а не только ДНК.
Вебер (2005) бросает вызов телеосемантике Мэйнарда Смита (2000). учетная запись. Мейнард Смит проводит аналогию между информацией в запрограммированный компьютер и информация в ДНК. Компьютеры выполняют алгоритмы, запрограммированные людьми и организмами, выражают ДНК, которая был запрограммирован естественным отбором. Информация запрограммирована в компьютере преднамеренно, чтобы можно было определить намерения человека-программиста путем анализа алгоритма.Мэйнард Смит утверждает, что информация, запрограммированная в ДНК естественным отбор намерен в том же смысле. Вебер предлагает два аргумента против этой точки зрения. Во-первых, он указывает, что ДНК может содержать нуклеотидные последовательности, возникшие в результате случайных мутаций, которые оказывается полезным. Если естественный отбор еще не сработал то телеосемантическая теория Мейнарда Смита подразумевает, что они не содержат информацию. Тем не менее, причинно такая нуклеотидная последовательность могла бы влияют на развитие так же, как и последовательности, которые были выбран для.Вторая критика Вебером счета Мейнарда Смита проистекает из более тщательного изучения интенциональности, связанной с компьютерные программы. Вебер утверждает, что намеренные состояния ассоциировали с компьютерами на самом деле являются состояниями инженеров-людей, которые пишут программы, а не состояния самих компьютеров: “Компьютер программа представляет собой строку символов, которая обретает смысл только в контекст сообщества инженеров, которые понимают, что программа делает и для чего его можно использовать »(Weber 2005, p.252). Аналог программистам в аккаунте Мейнарда Смита естественно выбор. Но естественный отбор не имеет преднамеренного состояния. Следовательно, заключает Вебер, телеосемантический подход не может сохранить идею о том, что ДНК содержит информацию в преднамеренном смысл.
Заманчиво думать, что информационный разговор в этом вопросе бессилен. контекст, и действительно, некоторые философы утверждали, что такие разговоры вводит в заблуждение и от него следует отказаться (например, Sarkar 1996, Weber 2005 и, возможно, Rosenberg 2006).Но другие взяли считают, что более внимательное отношение к концепциям информации может приводят к важным выводам (см. следующий раздел).
5.2 Использование широко биологической концепции информации для объяснения того, что делают гены
Цель Jablonka – дать общее определение информации. который распознает различные типы информации, связанной с различные способы получения, копирования и передачи информации сквозь пространство и время (Яблонька 2002). Одна из ее проблем заключается в том, что дискуссии о значении (или несмысле) информации говорят в биология склоняется к предположению, что генетическая система должна служат прототипом для размышлений о биологической информации.Она считает, что общее определение информации, предназначенное для улавливать смыслы информации, воплощенные в экологических подсказках, рукотворные инструкции и эволюционные биологические сигналы, а также чувство информации в наследственном материале, приведет к более полезным обобщения и перспективы.
Яблонька говорит, что чувство информации во всех этих ситуациях задействовать источник, принимающую систему (организм или созданный организмом системы), так и особого типа реакции приемника на источник.Она воспринимает реакцию получателя как сложную, регулируемую цепочка событий, приводящая к ответу. Вариации в форме источник приводят к вариациям в ответе. То есть характер реакция зависит от того, как организован источник. Кроме того, она указывает на то, что реакции в этих ситуациях полезны для приемник в течение соответствующего периода времени (в случае организмов, за эволюционное время). Яблонька подчеркивает, что выгода или функцию, в случае организмов следует понимать с точки зрения эволюция, с акцентом на эволюцию реакции системе , а не об эволюции источника или эволюции окончательный результат реакции.
Концепция информации Яблоньки является преднамеренной и связана с телеосемантические концепции, рассмотренные выше. По стандарту телеосемантические концепции, сигналы имеют информацию, потому что производство сигнала было выбрано в эволюционной истории. Однако, по мнению Яблоньки, у субъекта есть информация, а не потому что он был выбран для, но потому что ответ получателя на него был выбран для. Считается ли что-либо информацией, зависит от реагируют ли сущности на это (надлежащим) функциональным образом.
Яблонька резюмирует свое общее мнение следующим образом. определение:
Можно сказать, что источник – объект или процесс – имеет информация, когда система-приемник реагирует на этот источник в специальном способ. Реакция приемника на источник должна быть такой, чтобы реакция может фактически или потенциально изменить состояние приемник в (обычно) функциональной манере. Более того, должен быть согласованная связь между вариациями в форме источника и соответствующие изменения в ресивере.(Яблонька 2002, с. 582)
Яблонька отмечает, что в соответствии с этим определением гены не иметь теоретически привилегированный статус; они являются одним из многих источников информации. Кроме того, она настаивает на том, чтобы сосредоточить внимание на «Интерпретирующая система получателя информация », а не об источнике.
Яблонька утверждает, что информация в ДНК имеет мало общего с информация в сигнале тревоги, облачном небе или химическом сигнале в бактериальной колонии.В последних случаях приемники реакции (или «ответы») на источник адаптивны для приемник: «тревога предупреждает птицу, что есть хищники. около; облачное небо предупреждает обезьяну о приближающейся буре; в химикат предупреждает бактерии о неминуемой голодной смерти ». (стр.585). Но в случае с ДНК приемник вроде не реагирует таким образом, чтобы клетка адаптировалась ко всему конкретному. «Скорее, ДНК просто« считывается »клеткой, поэтому она не информация в том же смысле … ДНК – это информация “О” клетке или организме, а не «Для» клетки или организма.»(Яблонька 2002, п. 585). Тем не менее, Яблонька утверждает, что ее концепция применима к гены, даже если это не относится к ДНК в целом:
Однако если вместо того, чтобы думать о ДНК в целом, мы думаем о конкретный локус с определенным аллелем, несложно подумайте о функциональной роли этого конкретного аллеля в конкретный набор экологических обстоятельств. Следовательно, мы можем сказать, что для все типы информации , в том числе тревожные звонки и фрагменты ДНК, источник S (аллель, сигнал тревоги, пасмурное небо и др.) несет информация о состоянии E для приемника R ( организм или продукт, созданный для организма), если получатель имеет система интерпретации, которая реагирует на S так, как обычно заканчивает тем, что адаптирует R (или его разработчик, если R по-человечески спроектировано) по E . (Яблонька 2002, с. 585, мой стресс)
Учитывая, что Яблонька говорит, что ДНК вообще не является информацией в тот же смысл, что и звонок будильника и пасмурное небо (и что это смысл, указанный в заявлении выше), вызывает недоумение, почему она утверждает что приведенное выше утверждение относится ко «всем типам Информация.Кроме того, она утверждает, что приведенное выше утверждение относится к конкретным аллелям (и, по-видимому, не к ДНК в целом) не однозначно. В оригинальном аккаунте Яблоньки есть поучительный способ думать об информации в биологических процессах такие как клеточные сигнальные процессы. Но ее счет не обосновать идею о том, что гены и ДНК содержат информацию или помогают выяснить роль генов и ДНК.
5.3 Причинная интерпретация утверждений о том, что гены делают
Другой подход к выяснению роли генов и ДНК состоит в том, чтобы замените бессвязный информационный разговор конкретными причинными описаниями основаны на четком понимании причинно-следственной связи (Waters 2000, и готовятся к печати).Такой подход основан на идее, что базовый теория и лабораторные методы, связанные с молекулярной генетикой, могут быть понимается с чисто причинной точки зрения. Базовый теория и методология касается синтеза ДНК, РНК и молекулы полипептидов, а не предполагаемая роль ДНК в “программировании” или «руководящее» развитие (раздел 2.3). Причинная роль молекулярного гены в синтезе этих молекул можно понять в терминах причинно-следственных фактических различий.Это включает в себя два причинные концепции, фактические различия и причинные специфичность. Эти концепции могут быть объяснены в терминах возможность манипулирования причинно-следственной связью .
Концепция реального внесения изменений применяется в контексте фактическая совокупность, содержащая сущности, которые фактически отличаются от уважение к некоторой собственности. В такой популяции может быть много потенциалов производителей разницы. То есть может быть много факторы, которыми можно манипулировать, чтобы изменить соответствующее свойство сущности в населении.Но фактическая разница производители – это (грубо говоря) производители разности потенциалов, которые фактические различаются, и чьи фактические различия приводят к фактическим различия в собственности у населения.
Концепцию реального создания различий можно проиллюстрировать с помощью Принцип различия классической генетики (раздел 2.1). В соответствии с согласно этому принципу, гены могут создавать различия в отношении фенотипические различия в конкретных генетических и экологических контексты.Таким образом, он определяет потенциальных производителей разницы. Когда это принцип используется для объяснения фактического наследственного паттерна, это применяется к генам, которые действительно различались в популяции, проявляющей образец (часто экспериментальная популяция). В таких случаях фактическая разница в гене среди организмов в популяции вызвали фактические фенотипические различия в этой популяции (см. Гиффорд 1990). То есть ген был актуальным производитель разницы, а не просто производитель разницы потенциалов (в том численность населения).
Концепция реального изменения может быть применена к молекулярным генетика следующим образом. В реальной ячейке, где население молекулы необработанной РНК различаются по линейной последовательности, Возникает вопрос: чем вызваны эти различия? Ответ в том, что различия в генах в клетке вызывают фактические различия в линейные последовательности в необработанных молекулах РНК, а также в популяции молекул РНК и полипептидов. Гены не единственные фактические разницы, создающие фактические различия в линейных последовательности этих молекул.И это подводит нас ко второй причинной понятие, причинная специфика.
Причинная специфичность была проанализирована Льюисом (2000). Основная мысль состоит в том, что причинно-следственная связь между двумя свойствами специфична, когда много разных значений причинного свойства вызывают много разных и, в частности, разные значения результирующей переменной (причинная отношения создают что-то вроде математической функции). An переключатель включения / выключения не является специфическим в этом техническом смысле, потому что причинное свойство имеет только два значения (включено и выключено).Переключатель диммера причинно-специфический в этом смысле. Гены могут иметь конкретное различие производителей, потому что многие специфические различия в последовательностях нуклеотиды в ДНК приводят к специфическим различиям в РНК молекулы. Это не относится ко многим другим фактическим различиям. производители, такие как полимеразы, которые больше похожи на выключатели (с относительно различий в линейных последовательностях). Биологи обнаружил, однако, существование других фактических создателей различий, помимо генов и ДНК, которые причинно специфичны в отношении линейные последовательности процессированных РНК и полипептидов, в некоторой степени наименее.Например, в некоторых клетках сплайсинговые комплексы называются спликосомы на самом деле различаются множеством способов, что приводит к множеству, специфические различия в линейных последовательностях процессированной РНК молекулы.
Тот факт, что все виды сущностей причинно связаны с синтез РНК может привести к мысли, что существует причинная связь между причинные элементы. Но этот отчет показывает, что гены и ДНК играют отличительная причинная роль в том, что гены являются причинно-специфическими фактическими различия, которые создают различия в линейных последовательностях необработанных Молекулы РНК.Эта отличительная роль расширяется (с важными квалификации) к линейным последовательностям процессированных молекул РНК и полипептиды. Основная теория, связанная с молекулярной генетикой, так как отличается от фундаментальной теории, может быть объяснен в причинно-следственных связях. термины.
5.4 Причинно-следственные связи развития программ ДНК
Вебер (2005) и Розенберг (2006) независимо друг от друга утверждают, что ДНК содержит генетическую программу, которая выполняется в процессе разработки, но оба отрицают, что эта идея зависит от представления о том, что ДНК содержит семантические или преднамеренная информация.Они иллюстрируют этот момент, рассматривая современное объяснение первых стадий переднезаднего формирование паттерна у эмбрионов Drosophila . Это объяснение объясняет, как одноклеточный эмбрион с внутриклеточным градиентом бикоидного белка (концентрация бикоидного белка уменьшается от перед задним концом) развивается в многоклеточный зародыш с 14 парасегментов (первые три парасегмента позже образуют голову, следующие три сформируют грудную клетку, а остальные сегменты сформируют брюшные сегменты).Вебер начинает с объяснения того, что бикоид градиент «генерируется синтезом белка из мРНК. вид, который произошел от матери (путем транскрипции материнского гены) и откладывается в яйцеклетке на переднем конце материнские медсестры “. (Weber 2005, p. 244). объясняя, как белок-биоид по-разному активирует набор шесть пробелов генов, которые, в свою очередь, дифференцированно активируют и деактивировать восемь парных правил генов, которые дифференцированно активируют и деактивировать сегментов полярности генов и гомеотических селектор генов.
Детали каскадов активаций и деактиваций генов были исследованы экспериментально. Вебера и Розенберга резюме результатов показывают, что биологи могут объяснить начальные формирование картины с точки зрения последовательных градиентов концентрации, которые регулируют гены без существенного обращения к информационным концепциям. Вебер подчеркивает этот момент, подробно утверждая, что использование термина «Позиционная информация» является метафорической, и что действие бикоидного градиента можно объяснить без концепции Информация.Он приходит к выводу, что этот градиент и другие не несут информация в преднамеренном смысле; скорее, они играют причинную роль в формирование новых градиентов через регуляцию генов.
Розенберг обсуждает проблемы, противостоящие идее о том, что ДНК содержит генетическая программа развития. Он начинает с вопроса о учитывается ли ранняя стадия развития паттернов, которая он и Вебер резюмируют (по отдельности), могут быть развиты, чтобы дать «вся история развития» (Розенберг 2006, стр.75). Будет ли весь история быть макромолекулярной? Он также указывает, что объяснение Формирование паттернов, которые он описывает, сформулировано в терминах генов. Он спрашивает: Выживет ли это объяснение, если не исчезнет концепция гена? Но в в конце, он выражает оптимизм в великой идее о том, что объяснение начальная разработка модели может быть разработана для учета весь процесс развития.
Хотя и Розенберг, и Вебер отвергают идею о том, что биологи объяснение начальной стадии формирования паттерна делает существенное ссылка на идею, что гены или ДНК имеют преднамеренную информацию, они предполагают, что объяснение содержит существенную ссылку или подразумевает наличие генетической программы.Но неясно, почему использование термин программа менее метафоричен, чем термин позиционная информация . Розенберг утверждает, что мы можем сделать вывод что ДНК может выполнять программы, потому что компьютеры могут быть построены из ДНК, и эти компьютеры на основе ДНК могут выполнять программы в в том же смысле, что компьютеры на основе кремниевых чипов могут выполнять программы. Тем не менее, непонятно, к чему добавляется фраза «выполнение генетической программы» дополняет объяснение начального формирование рисунка.Вебер и Розенберг представляют свои резюме объяснение в терминах причинности, таких как «генерировать», “Депозит”, “активировать” и «Деактивирован». Можно спросить, действительно ли причинное объяснение заменяет метафорическую идею о том, что ДНК программирует развитие.
5.5 Фундаментальное теоретизирование против исследовательской прагматики
Учетные записи, подобные представленным выше, в случае успеха предоставляют лучшая основа для понимания базовой теории связанного с молекулярной генетикой, но неясно, могут ли они прояснить идея о том, что гены – это «фундаментальные» сущности, которые «программируют» развитие и функционирование организмов путем “управления” синтезами белков, которые, в свою очередь, регулируют все важные клеточные процессы.На самом деле, есть значительный скептицизм в отношении философское сообщество об этой фундаментальной теории. Обычный критика (среди философов) состоит в том, что она ориентирована на гены.
Некоторые философы решили заменить фундаментальную теорию связаны с молекулярной генетикой с новой фундаментальной теорией, которая не делает ген “привилегированным”. Среди предложений Роберта (2004). Опираясь на труды Буриана, Гриффитса, Келлера, Оямы, Мосс и другие, Роберт пытается построить новую «основу для понимание и объяснение развития организма »(стр.78) что не сводит «проблему развития» к проблема действия гена и активации гена. Он ищет рамки, которые свободна от предполагаемого предположения, что развитие включает в себя выражение «заранее сформированной генетической информации» (стр. 56). Роберт говорит, что его концепция фокусируется на организмах, а не на гены и что он «серьезно» Келлер подчеркивает сложности развития.
Философы, занимающиеся фундаментальным теоретизированием, по крайней мере, еще не привлекла большое внимание практикующих ученых.Возможно отсутствие внимания связано с разницей между проблемами (многих) философы биологии и потребности практики биологи. Вагнер, биолог-теоретик, проводивший математическая / концептуальная работа в эволюционной теории, а также эмпирическое исследование эволюции морфологических признаков, делает следующее наблюдение в обзоре книги Роберта,
Роберт и его коллеги, которые выступают за аналогичную интерпретацию, правильным в том смысле, что сами по себе гены не могут создать организм, а вместо этого встроены в большую сеть причинно-следственных связей.Но ученые обычно не интересуются общими утверждениями о том, что в принципе является требуется, чтобы понять явление.
Да, ученые принимают прагматичное решение о том, что изучать, но я думаю, что эти решения отнюдь не произвольны. Сила молекулярно-генетические подходы дались нелегко, скорее это результат долгой истории напряженных исследований, основанных на видении, которое получено из работ Ричарда Гольдшмидта, Альфреда Кюн и Томас Хант Морган в начале 20 века.Таким образом, генетическими исследованиями движет не интеллектуальная лень. программа; вместо этого мы загребаем трофеи от с трудом одержанной победы над биологическая сложность. (Вагнер 2004, с. 1405)
Несоответствие между интересами (многих) философов к фундаментальные теоретические и практические биологи потребность сделать прагматические решения о том, что изучать, поднимает важный вопрос о науках, ориентированных на гены, которые в значительной степени игнорировались философы: почему так много биологических исследований сосредоточено на генах и ДНК?
В официальном и публичном контексте ученые обращаются к фундаментальным теория, связанная с молекулярной генетикой, чтобы оправдать центрирование исследования генов и ДНК (например,ж., см. сайты финансирующих агентств например, Национальный центр биотехнологической информации). Гены обычно называемые «фундаментальными единицами», которые отвечает за руководство всеми основными жизненными процессами. Обычно сочетание причинно-следственных и информационных метафор используется для объяснения роль генов. Говорят, что гены производят РНК и полипептиды, чтобы предоставлять инструкции или управлять процессами. Но философский расследование показало, что такого рода широкие претензии не могут выдерживают тщательное изучение.Почему же тогда так много исследований сосредоточено на гены и ДНК? Один из ответов на этот вопрос заключается в том, что биологи ослепленные идеологией генетического детерминизма. Но защита Вагнера геноцентризм предлагает другой ответ, ответ, который резонирует с Объяснение Келлера (2000) того, почему разговор о генах полезен.
Было высказано предположение, что настоящая причина, по которой биологи привлекают внимание на генах и ДНК заключается в том, что гены создают различия, которые можно использовать для отслеживания и управления широким спектром биологических процессов (воды 2004a и 2006).Эта научная практика имеет смысл независимо от любая фундаментальная теория, связанная с молекулярной генетикой. В этом случае молекулярной генетики, это исследовательская прагматика, а не фундаментальная теоретизирование, которое движет научными исследованиями. Достаточно базовой теории для объяснения исследовательской полезности и результатов генно-ориентированных подходы. Фундаментальная теория в важном смысле эпифеноменальный в отношении дизайна и реализации геноцентрические исследования. С этой точки зрения роль фундаментальных теорию следует понимать в терминах Латура (1987, 1988), как платформа для сплочения войск и привлечения ресурсов на исследования усилия.План лабораторных экспериментов и причина почему эксперименты работают, можно объяснить с точки зрения общих исследовательские стратегии, основная каузальная теория молекулярных генетика и детали экспериментальных контекстов.
Философия молекулярной генетики – это оживленная область исследований, которая отражает большую часть волнения и разнообразия современного философия науки. Философы, исследующие смежные области генетики имеют разные философские интересы и принимают противоположные подходы.Некоторые философские исследования направлены на установление науки прямо, в то время как другие исследования направлены на исправление нашего понимания науки. Некоторые философы используют традиционные методы философии. науки, например, анализ концепций и моделей, в то время как другие открывают новые философские подходы, опираясь на методы из социальных наук, включая анкетные исследования. Хотя некоторые философы сосредотачивают свое внимание на фундаментальном теоретизировании, другие исследуют динамику эмпирических исследований.Исследования в философия молекулярной генетики также генерирует новые идеи об основных концепциях, представляющих общефилософский интерес, включая редукционизм, информация и причинно-следственная связь.
Классическая генетика
Классическая генетикаКлассическая генетика основана на делении клеток с участием хромосом, которые состоят из единиц наследования, называемых генами. Соматические клетки, содержащие 46 хромосом (22 пары автомосом и две половые хромосомы, XX или YY) делятся путем митоза.При митозе в одной соматической клетке образуются две дочерние клетки, каждая с диплоидным числом хромосом (46). Мейоз происходит только в половых клетках и дает две гаметы с гаплоидным числом хромосом каждая (23). Подробное описание митоза и мейоза см. В любом стандартном тексте по генетике в разделе «Рекомендуемая литература».
Гены хромосом подчиняются законам Менделя следующим образом:
- Наследование основано на «факторах» (генах), которые передаются потомству как дискретные единицы.Для каждого признака у человека есть два фактора (аллеля), по одному от каждого родителя.
- Два аллеля всегда разделяются и переходят к разным гаметам.
- Факторы для разных признаков распределяются по гаметам независимо, т.е. материнские и отцовские гены случайным образом рекомбинируют в гаметах.
За прошедшие годы в законы Менделя были внесены некоторые уточнения. Поскольку тесно связанные гены на одной хромосоме не сортируются независимо друг от друга, они всегда наследуются вместе.Напротив, гены, локусы которых находятся далеко друг от друга на хромосоме, могут испытывать кроссинговер, при котором у потомства присутствует новая комбинация генов (рекомбинантная). Кроме того, нерасхождение может происходить (редкое событие), когда две хромосомы не могут разделиться во время мейоза, так что обе передаются одной дочерней клетке.
Принципы классической генетики могут быть применены к наследованию генов группы крови . Для иллюстрации: для системы групп крови ABO родители AO и BO могут производить потомство всех групп ABO (рис. 1-1).
Вместо нормальной сегрегации может возникнуть нерасхождение , хотя и редко (рис. 1-2).
Для тесно связанных генов на хромосоме система групп крови Rh служит моделью. Поскольку локусы Dd, Cc и Ee тесно сцеплены на хромосоме, кроссинговера не происходит (рис. 1-3).
Чтобы проиллюстрировать переход между и , полезна система групп крови MNS (рис. 1-4). В этой системе M и N являются аллелями в одном локусе, как S и s .Локус MN и локус Ss сцеплены, но не так тесно, как локусы в системе Rh.
В трансфузионной медицине мы используем эти принципы при исследовании случаев гемолитической болезни новорожденных, при исследовании того, являются ли недавно обнаруженные гены группы крови независимыми системами или являются частью существующей системы, а также при рассмотрении споров об отцовстве. Применение менделевской генетики к группам крови будет снова проиллюстрировано позже, когда будут обсуждаться некоторые из общих систем групп крови.
Деятельность по обогащению 1
Сегодня генетика продвинулась туда, где мы теперь можем клонировать овец, что привело к продолжающимся спорам о последствиях клонирования человека. Чтобы определить некоторые из проблем, прочитайте как минимум три из следующих кратких статей из British Medical Journal (BMJ) .
- Отправьте комментарий или вопрос в список рассылки класса по любой проблеме, поднятой в статьях BMJ .
- Также отправьте комментарий или ответ в список рассылки класса в ответ на комментарий или вопрос, сделанный другим участником.
Условия для поиска
- аллель
- кроссинговер
- диплоидное число хромосом
- гамета
- гаплоидное число хромосом
- сцепленное 9055 локус мейоз6 синтенический
| Историческая перспектива | Классическая генетика | Генетические символы |
Классическая менделевская генетика (закономерности наследования) – понимание генетики
Основные законы наследования важны для понимания паттернов болезни коробка передач.Паттерны наследования заболеваний с одним геном часто называют Менделирующие со времен Грегора Менделя впервые наблюдали различные модели сегрегации генов для отобранных признаков у огородного гороха и смог определить вероятность повторения черта для последующих поколений. Если семья поражена болезнью, верная семья история будет важна для установления модели передачи. Кроме того, семья анамнез может даже помочь исключить генетические заболевания, особенно распространенные заболевания, при которых поведение и окружающая среда играют важную роль.
Большинство генов имеют одну или несколько версий из-за мутаций или полиморфизмов, называемых аллели. Люди могут нести «нормальный» аллель и / или «Болезнь» или «редкий» аллель в зависимости от воздействия мутация / полиморфизм (например, болезнь или нейтральный) и популяционная частота аллеля. Заболевания с одним геном обычно наследуются по одной из нескольких схем в зависимости от расположение гена и необходимость одной или двух нормальных копий гена для проявление фенотипа болезни.
Экспрессия мутировавшего аллеля по отношению к нормальному аллелю может быть охарактеризована как доминантный, содоминантный или рецессивный. Существует пять основных режимов наследования для моногенные заболевания: аутосомно-доминантный, аутосомно-рецессивный, X-связанный доминантный, X-связанный рецессивный и митохондриальный.
Генетическая гетерогенность – обычное явление как для моногенных заболеваний, так и для сложных многофакторные заболевания. Неудивительно, что несколько затронутых членов семьи могут иметь различную степень тяжести и исходов заболевания.Этот эффект может быть вызван другие гены, влияющие на фенотип заболевания, или разные мутации в том же гене приводя к сходным, но не идентичным фенотипам. Некоторые отличные ресурсы для информации о моногенных заболеваниях – это онлайн-Менделирующее наследование у человека (OMIM; http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?db=OMIM) и GeneTests / GeneClinics (http://www.genetests.org).
Паттерны наследования
Аутосомно-доминантный
Аутосомно-рецессивный
Х-сцепленный доминант
Х-сцепленный рецессивный
Митохондриальный грибок, как и самки передается только самками
Может появляться в каждом поколении
Просмотр в собственном окне
| Образец наследования | Примеры заболеваний |
|---|---|
| Аутосомно-доминантный | болезнь Хантингтона нейрофиброматоз, ахондроплазия, семейная гиперхолестеринемия |
| Аутосомно-рецессивный | Болезнь Тея-Сакса, серповидно-клеточная анемия, кистозная фиброз, фенилкетонурия (PKU) |
| Х-сцепленный доминантный | Гипофатемический рахит (резистентный к витамину D рахит), дефицит орнитин-транскарбамилазы |
| Х-сцепленный рецессивный | Гемофилия А, мышечная дистрофия Дюшенна |
| Митохондриальная | наследственная оптическая нейропатия Лебера, Синдром Кернса-Сейра |
От классической генетики к количественной генетике к системной биологии: моделирование эпистаза
Abstract
Данные по экспрессии генов использовались вместо фенотипа как в классических, так и в количественных генетических параметрах.Эти две дисциплины имеют разные подходы к измерению и интерпретации эпистаза, который представляет собой взаимодействие между аллелями в разных локусах. Мы предлагаем основу для оценки и интерпретации эпистаза на основе классического эксперимента, который сочетает в себе сильные стороны каждого подхода. Этап регрессионного анализа учитывает количественный характер измерений экспрессии, оценивая эффект делеций генов плюс любое взаимодействие. Эффекты выбираются по значимости, так что сокращенная модель описывает каждый признак выражения.Мы показываем, как полученные модели соответствуют конкретным иерархическим отношениям между двумя генами-регуляторами и целевым геном. Эти отношения являются основными единицами генетических путей и диаграмм геномной системы. Наш подход может быть расширен для анализа данных из различных экспериментов, нескольких локусов и различных сред.
Сведения об авторе
Эпистаз долгое время имел два немного разных значения в зависимости от контекста, в котором он обсуждается. Классическое определение описывает аллель в одном локусе, полностью маскируя эффект аллеля во втором локусе.Такие отношения можно интерпретировать как иерархические, и их можно комбинировать для определения генетических путей. В количественной генетике эпистаз охватывает широкий спектр взаимодействий и может распространяться более чем на два локуса. Эти два определения сосуществуют, потому что они обычно применяются к разным типам исследуемых групп и разным типам черт. Текущая тенденция заключается в том, чтобы рассматривать экспрессию генов как признак в различных генетических фонах. Это дает повод вернуться к эпистазу в этом новом контексте.Мы принимаем непрерывный характер экспрессии генов, используя идеи количественной генетики, но сохраняем иерархическую интерпретацию классического эксперимента. Эти иерархические отношения являются строительными блоками системных диаграмм и генетических путей. Эта структура может служить основой для будущего анализа эпистаза на основе геномных данных.
Образец цитирования: Aylor DL, Zeng Z-B (2008) От классической генетики до количественной генетики и системной биологии: моделирование эпистаза.PLoS Genet 4 (3): e1000029. https://doi.org/10.1371/journal.pgen.1000029
Редактор: Дэвид Б. Эллисон, Университет Алабамы в Бирмингеме, Соединенные Штаты Америки
Поступила: 12 июня 2007 г .; Принята к печати: 8 февраля 2008 г .; Опубликовано: 14 марта 2008 г.
Авторские права: © 2008 Aylor and Zeng. Это статья в открытом доступе, распространяемая в соответствии с условиями лицензии Creative Commons Attribution License, которая разрешает неограниченное использование, распространение и воспроизведение на любом носителе при условии указания автора и источника.
Финансирование: Эта работа была частично поддержана NIH GM45344 и Национальной исследовательской инициативой Государственной кооперативной службы исследований, образования и распространения знаний Министерства сельского хозяйства США, номер гранта 2005-00754. DLA поддерживается грантом NIEHS на обучение (T32 ES007329) в области биоинформатики.
Конкурирующие интересы: Авторы заявили об отсутствии конкурирующих интересов.
Введение
Эпистаз традиционно обсуждался в двух различных контекстах, соответствующих дисциплинам классической молекулярной генетики и количественной генетики.В каждом случае термин описывает взаимодействие между аллелями в двух или более локусах. Однако методы обнаружения эпистаза и интерпретации лежащей в основе биологии сохранили исторические разногласия, несмотря на призывы к синтезу [1]. Во многом это связано с тем, что в этих двух областях традиционно изучаются разные типы черт в разных экспериментальных популяциях.
Классический эксперимент с эпистазом сравнивает двойной мутант с двумя ассоциированными одиночными мутантами. Эпистаз присутствует, если наблюдаемый фенотип с двойным мутантом отнесен к той же категории, что и фенотип с одним мутантом.Это подразумевает особый тип взаимодействия, при котором аллель в одном локусе маскирует эффект вариации во втором локусе. Эта взаимосвязь описывается как эпистатический первый локус по отношению ко второму и может интерпретироваться как один ген, действующий выше другого. Эта иерархическая интерпретация была использована для построения биологических путей через серию пар эпистатических генов. Однако этот подход ограничен необходимостью легко наблюдаемых и категоризированных фенотипов [2].
Напротив, количественная генетика изучает признаки, которые постоянно меняются и не могут быть легко разделены на категории.Такое распределение признаков является результатом кумулятивного воздействия многих генов. Каждый дополнительный ген увеличивает возможную комбинацию аллелей, а количество возможных фенотипов растет в геометрической прогрессии. Фенотип человека – это сумма аллельных эффектов каждого гена и влияния окружающей среды. Эпистаз определяется как отклонение от этих аддитивных генных эффектов [3]. Количественная генетическая модель может включать множественные локусы и множественные взаимодействия. Эпистаз в этом смысле описывает функциональные отношения между генами в контексте признака, но он включает в себя как иерархические, так и неиерархические отношения, и нет способа провести различие между ними.
Любой генетический эффект актуален только для изучаемой популяции из-за наличия генетического фона. Фон – это генетическая изменчивость, которая не наблюдается в популяции и не может быть смоделирована. Классический эксперимент проводится с использованием генетически однородных лабораторных штаммов, поэтому фон отсутствует. Количественная генетика изучает различные популяции, и фоновые вариации почти всегда присутствуют. Это означает, что эпистаз может быть обнаружен в одном эксперименте, но не в другом.Это привело к критике, что эпистаз в количественном генетическом смысле является статистической конструкцией, а не истинным представлением биологии.
Фактически, оба подхода стремятся проиллюстрировать лежащую в основе молекулярную архитектуру, и каждый из них имеет свои сильные стороны. Иерархическая интерпретация эпистаза привлекательна, поскольку повышенное внимание уделяется генетическим путям и системным диаграммам. Однако количественные подходы необходимы для учета непрерывных типов данных, таких как экспрессия генов, концентрации метаболитов и пригодность.Недавняя литература предполагает, что такие подходы принимаются. Например, в то время как ранние крупномасштабные профили приспособленности у мутантов с делецией дрожжей [4], [5] оценивались категориально, St Onge et al [6] измерили приспособленность 650 штаммов дрожжей с двойной делецией и применили новый количественный анализ.
Развитие геномных методов сломало один из традиционных барьеров, обсуждавшихся выше: одни и те же черты теперь используются как в классических, так и в количественных параметрах [7]. Экспрессия генов, пожалуй, самый распространенный пример.Вместо одного значения фенотипического признака каждого человека описывает вектор измерений экспрессии. Профили экспрессии в штаммах дрожжей с одной делецией показали, что 34% мутантов показали двадцать или более дифференциально экспрессируемых генов [2]. При картировании локуса количественных признаков экспрессии (eQTL) используется подход линейного моделирования, чтобы связать генетические вариации с признаками экспрессии генов [8] – [12]; Стори и др. [13] обнаружили, что более тридцати процентов признаков были совместно связаны с двумя локусами у дрожжей. Когда экспрессия гена коррелирует со сложным фенотипом, соответствующие признаки могут отражать молекулярную основу этого признака на промежуточном уровне между генотипом и фенотипом.Некоторые исследования предполагают, что эпистаз широко распространен среди экспрессионных признаков [14] – [16], и такие признаки могут иметь больше QTL, чем классические признаки [13], [17]. Поскольку экспрессия генов используется как в классическом, так и в количественном контексте, это ценная основа для сравнения способности обнаруживать эпистаз и интерпретировать характер взаимоотношений между генами.
Мы предлагаем основу для оценки и интерпретации эпистаза с использованием признаков экспрессии. Наша цель – учесть непрерывный характер данных, но при этом сохранить иерархическую интерпретацию эпистаза.Такие интерпретации хорошо известны для классических экспериментов с эпистазом [18], но только недавно были изучены для сложных данных [19]. Мы уточняем классические интерпретации, явно моделируя экспрессию генов. Эффекты и взаимодействия генов оцениваются с использованием линейной модели, аналогично картированию eQTL. Наш метод выбирает наиболее подходящую регрессионную модель для каждого признака, которая описывает порядок и характер функции гена. Такие отношения являются основными единицами генетических путей и системной биологии.Мы специально обращаемся к тому, как использовать непрерывный фенотип, как статистически обоснованным, так и совместимым с классическим подходом.
Мы проиллюстрируем наш метод общедоступными измерениями экспрессии Dictyostellium discoideum дикого типа [20] и мутантных штаммов с делециями [21]. Этот эксперимент представляет собой классический анализ эпистаза, который нацелен на гены пути протеинкиназы (PKA) и измеряет профиль экспрессии генов каждого штамма.
Результаты
Моделирование эпистаза для постоянно изменяющихся черт
В классическом анализе эпистаза триплеты делеционных мутантов объединяются с диким типом, чтобы сформировать контраст.Каждый контраст включает два одиночных мутанта и двойной мутант. Каждый описан относительно известного фенотипа дикого типа. Гипотетический пример признака, на который влияют два гена, A и B , можно описать следующим образом, где y – значение признака, μ – ожидаемое значение дикого типа, β A и β B – это эффекты удаления каждого гена, а ε – член ошибки.
Это строго соответствует классическому определению, но есть явная проблема; нет положения, если двойной мутант не попадает точно в ту же категорию, что и один из одиночных мутантов.По этой причине признаки экспрессии генов плохо вписываются в классические рамки. Выражение непрерывное, ожидаются промежуточные уровни. Более того, даже нормализованные значения признаков неизбежно будут включать некоторую погрешность измерения. По этим причинам наблюдение двойного мутанта редко совпадает с наблюдением одного мутанта или наблюдения дикого типа. Предыдущие исследования пытались обойти эту проблему, полагаясь на различия между мутантами, чтобы определить наиболее похожую пару мутантов.Однако предположение о том, что выражение лица полностью замаскировано, неверно. Чтобы решить эти проблемы, мы уходим от прямого сравнения значений характеристик. Вместо этого мы оцениваем каждую делецию в зависимости от того, существенно ли она влияет на экспрессию мишени, и связываем уникальные паттерны значимости с моделями действия генов.
Мы используем линейную модель для оценки эффекта каждого удаления. Это общий способ связать всех мутантов с диким типом без каких-либо предположений о природе двойного мутанта.Мы регрессируем значение признака (например, выражение) на индикаторных переменных, представляющих наличие или отсутствие каждого аллеля дикого типа и члена взаимодействия. Взаимодействие описывает эффекты, уникальные для двойного мутанта. Тот же пример, рассмотренный выше, можно описать следующим образом.
Значение признака = Дикий тип + Эффект удаления A + Эффект удаления B + Взаимодействие + ошибка
Для построения такой линейной модели можно использовать различные методы. Сначала мы подбираем полную модель, а затем используем пошаговый выбор в обратном порядке, чтобы отбросить члены модели с коэффициентами, которые не имеют значения на заданном уровне.Полученная сокращенная модель называется моделью наилучшего соответствия. Для любого признака существует восемь возможных моделей, наиболее подходящих для вас. Для наглядности пронумеруем уменьшенные модели следующим образом:
После определения наиболее подходящей модели мы оцениваем значения параметров, используя эту модель для каждого признака. Таким образом, у нас есть наиболее подходящая модель и оценки коэффициентов для каждого признака. Термины в каждой наиболее подходящей модели представляют собой значимые эффекты гена и взаимодействия, воздействующие на этот признак. Индивидуальные коэффициенты представляют собой предполагаемый эффект удаления каждого гена.Модель 7 соответствует классической модели выше, когда взаимодействие между двумя делециями компенсирует эффект одной из них, либо β I = – β A или β I = – β В . Модель 8 описывает случай, когда удаленные локусы не влияют на признак.
Модель наилучшего соответствия описывает каждый признак экспрессии гена. Таким образом, мы имели дело с проблемой непрерывной переменной. Однако, приняв количественную генетическую модель, мы потеряли привлекательную особенность классического эксперимента: способность интерпретировать иерархические отношения.В следующем разделе мы идентифицируем шестнадцать иерархических отношений и предлагаем, чтобы конкретная наиболее подходящая модель поддерживала каждое из них.
Интерпретация иерархического эпистаза
В количественной генетике термин взаимодействия в вышеупомянутой модели считается эпистазом. Однако эпистаз в этом смысле включает как иерархические, так и неиерархические отношения. И наоборот, хотя Модель 7 можно четко интерпретировать как иерархический эпистаз с условиями, описанными выше, она не применима ко всем возможным иерархиям.
Мы рассмотрели все комбинации порядка и действия генов в рамках простых моделей включения / выключения, а затем предсказали гипотетический эффект удаления генов на каждый из них (рисунок 1, рисунки S1, S2 и S3). Есть четыре точки вариации для моделирования для каждой пары генов. Первый – это идентичность вышестоящего гена, то есть порядок гена. Во-вторых, вышестоящий ген будет либо включать (усиливать), либо выключать (подавлять) нижележащий ген. В-третьих, нижележащий ген может усиливать или подавлять экспрессию целевого гена, экспрессия которого наблюдается.Наконец, мы считаем, что сам вышележащий ген будет усилен или подавлен некоторым инициирующим фактором, таким как сигнал развития или возмущение окружающей среды. Эйвери и Вассерман [18] предоставляют общую структуру, которая широко используется для интерпретации эпистаза в ответ на такие сигналы, и отмечают, что эффект мутации наблюдается только для определенного состояния сигнала. Однако знание состояния сигнала не дает никакой информации о том, усиливается или репрессируется вышестоящий ген в этом состоянии.В наших моделях мы фокусируемся на влиянии на вышестоящий ген. Эта модель имеет шестнадцать возможных вариантов, описывающих иерархические отношения между двумя генами и целевым геном.
Рис. 1. Моделирование взаимосвязи A является вышестоящим репрессором B .
B , в свою очередь, усиливает целевой ген X . В этом примере удаление A изменит состояние целевого гена с «выключено» на «включено». Поэтому мы включаем эффект A в соответствующую регрессионную модель.Удаление B оставляет целевой ген в том же состоянии, что и дикий тип, и его эффект не учитывается. Также не ожидается, что двойной мутант AB будет отличаться от дикого типа, несмотря на значимость делеции A . Поскольку эффект A уже включен в модель для этого контраста, он должен быть компенсирован членом взаимодействия. Мы заключаем, что если A усиливается сигналом, A репрессирует B , а B увеличивает X , соответствующая модель регрессии наилучшего соответствия будет включать коэффициенты для A и член взаимодействия.Аналогичная логика применима к случаю, когда сигнал подавляет A . Сигнал подавляет A , таким образом, удаление A не оказывает никакого влияния на нисходящий поток. Мы ожидаем только коэффициента, соответствующего нижележащему гену в наиболее подходящей модели.
https://doi.org/10.1371/journal.pgen.1000029.g001
Ключ к нашему подходу – соединение каждой из шестнадцати иерархических моделей с одной из восьми возможных наиболее подходящих регрессионных моделей. Если делеция изменяет состояние целевого гена относительно дикого типа у мутанта, то предполагается, что эта делеция окажет значительный эффект, и она будет включена в регрессионную модель, соответствующую этой иерархической модели.На рисунке 1 приведен пример одной из возможных моделей, в которой A усилен сигналом; A – репрессор перед B ; и B усиливает целевой ген X . Мы заключаем, что соответствующая модель регрессии наилучшего соответствия будет включать коэффициенты для A и член взаимодействия. Обратите внимание, что если сигнал вместо этого воспроизводит A , другая модель наилучшего соответствия представляет те же отношения между A и B .
Мы применили один и тот же подход к каждому из шестнадцати случаев и отметили несколько тенденций. Во-первых, влияние нижележащего гена на целевой ген X не влияет на соответствующую модель наилучшего соответствия. Это позволяет сократить пространство модели до восьми иерархических отношений (Таблица 1). Это наблюдение удобно, поскольку признаки экспрессии представляют все гены, расположенные ниже по ходу делеции. Независимо от прямого воздействия нижележащего гена, некоторые черты будут усилены, а другие подавлены.Когда вышестоящий ген является репрессором, четыре различных регрессионных модели представляют четыре уникальных иерархических отношения. Мы можем однозначно идентифицировать как порядок генов, так и влияние сигнала на вышестоящий ген. Мы не можем различить порядок генов, если вышестоящий ген является энхансером, потому что одна и та же наиболее подходящая модель описывает обе иерархии. Если вышестоящий ген просто усиливает действие нижележащего гена, удаление любого гена повлияет на ген признака аналогичным образом. Шесть из восьми возможных моделей регрессии наилучшего соответствия соответствуют восьми иерархическим отношениям.Примечательно, что иерархии можно указывать даже без эффекта взаимодействия в модели.
Мы также должны учитывать, что нет никакой иерархической связи между A и B , или что они не влияют на целевой ген (Таблица 1). Мы можем различать два типа параллелизма. Модель 4, двухгенная аддитивная модель без взаимодействия, не представляет собой эпистаза. Модель 3 представляет собой буферный эпистаз, при котором оба гена действуют на мишень в одном и том же направлении, и эффект удаления любого из них не очевиден, если оба гена не удалены.Мы называем это неиерархическим эпистазом, поскольку ни один ген не находится выше другого. Удаление деактивированного гена-регулятора не влияет на ген-мишень, что делает невозможным определение биологической взаимосвязи при деактивации регуляторов.
Остальная часть таблицы 1 представляет случаи, в которых один или оба гена не влияют на целевой ген. Признаки экспрессии, поддерживающие Модель 8 (без значимых терминов), могут представлять гены-мишени, которые не лежат ниже A или B и являются неинформативными.Результатом являются отношения один-ко-многим между наиболее подходящими моделями регрессии 1, 2 и 8 и соответствующими моделями экспрессии генов. Если вышестоящий ген иерархической пары отключен, мы не можем знать, является ли он вышестоящим или не задействованным.
Обычно экспрессия измеряется тысячами генов одновременно, и мы не ожидаем, что все они будут информативными. Даже при четкой интерпретации каждой характеристики в отдельности возникает проблема, связанная с интерпретацией всех черт вместе. Мы исследуем распределение всех признаков.Среди информативных признаков, связанных с наиболее подходящей моделью, большинство может представлять лежащие в основе биологические отношения между удаленными генами.
Проверка основы двухэтапного моделирования
Van Driessche et al. использовали Dictyostellium discoideum дикого типа [20] и мутантные штаммы с делециями [21], чтобы сделать вывод об иерархическом эпистазе среди генов пути протеинкиназы (PKA). Профиль экспрессии гена каждого штамма измеряли с использованием микрочипов кДНК с общим эталоном в течение 24 часов.Эти данные хорошо подходят для тестирования наших методов по двум причинам. Во-первых, эпистатические отношения между удаленными генами уже охарактеризованы экспериментально. Во-вторых, мутантные штаммы генетически идентичны по всем локусам, за исключением нескольких изучаемых, т. Е. Их генетический фон не отличается.
Путь PKA связан с реакцией агрегации развития на депривацию питательных веществ, которая инициируется на полпути во времени. Данные до и после агрегирования рассматривались отдельно, поэтому мы можем четко интерпретировать эффекты удаления в каждом состоянии сигнала.Данные представляли кратное изменение в логарифмической шкале, что делало распределение измерений экспрессии приблизительно нормальным; мы рассматриваем последствия этого в ходе обсуждения. Мы изучили 1553 признака экспрессии. Используемые нами гены были измерены в обоих экспериментах и по-разному экспрессировались в организме дикого типа во время агрегации [20]. Пять делеционных штаммов нацелены на гены пути протеинкиназы A (PKA), который участвует в ответе на голодание и активирует агрегацию. Это обеспечило три контраста: pufA / pkaC , pufA / yakA и regA / pkaR .Хотя существует десять возможных контрастов для этих пяти генов, были созданы только эти три двойных мутанта, предположительно потому, что они известны прямым родством.
Для каждого контраста некоторые черты поддерживали каждую модель (рис. 2). Кроме того, большое количество признаков не показало никаких эффектов удаления (т.е.поддержка модели 8). При пороге значимости p <0,01 большинство признаков поддерживали Модель 8 для каждой предварительной агрегации контраста (рис. S4) и для постагрегации контраста regA / pkaR .Согласно нашим интерпретирующим моделям, Модель 8 может указывать на три возможности. Первые два – это иерархические отношения, в которых вышестоящий усиливающий ген отключается во время агрегации. Последняя возможность состоит в том, что гены не участвуют в экспрессии мишени, и делеции не имеют никакого эффекта.
Рис. 2. Распределение наиболее подходящих моделей после агрегирования при пороге значимости p <0,01.
Частотное распределение наиболее подходящих регрессионных моделей можно интерпретировать как иерархические отношения между генами.Модель 8 соответствует отсутствию эффектов удаления и поддерживается большим количеством признаков в каждом контрасте; эти гены, вероятно, не находятся ниже по течению от делеций. Предполагается, что модель, поддерживаемая большинством оставшихся признаков, представляет истинные отношения.
https://doi.org/10.1371/journal.pgen.1000029.g002
Поскольку не все гены-мишени находятся ниже по ходу пути PKA, логично, что делеции не влияют на эти гены. Точно так же путь PKA запускается во время агрегации, и из этого следует, что делеции могут влиять на экспрессию только после того, как агрегация началась.Мы предполагаем, что гены-мишени, поддерживающие Модель 8, не находятся ниже по ходу пути, и что большинство оставшихся генов-мишеней отражают взаимосвязь внутри пути. Чтобы проверить это предположение, мы рассмотрели перекрытие между признаками экспрессии, поддерживающими Модель 8, для каждого контраста. Мы обнаружили, что все признаки выражения, поддерживающие модель 8 для контраста pufA / yakA , также поддерживали модель 8 для двух других контрастов. Эти черты убедительно подтверждают предположение, что они не находятся ниже по ходу пути PKA.
Когда мы посмотрели на эти гены для контрастов pufA / pkaC и pufA / yakA , одна модель получила сильную поддержку по сравнению со всеми остальными пост-агрегационными. Эти модели не только объясняют больше признаков после агрегирования, но и лучше подходят. В среднем наиболее подходящая модель объясняет более половины вариации выражения (R 2 ≥0,5, с поправкой на степени свободы в модели) для признаков в pufA / pkaC и pufA / yakA контрастов, и для обоих контрастов R 2 увеличился после агрегации (t-тест с p <0.0001).
Для контраста pufA / pkaC модель 2 имела наибольшую поддержку из семи ненулевых моделей. Модель 2 соответствует двум возможным интерпретациям. Во-первых, pkaC является нижележащим геном, что pufA является репрессором и что pufA отключается при наличии сигнала агрегации. С другой стороны, мы могли бы интерпретировать это как означающее, что только pkaC оказывает влияние на последующие цели и что pufA не имеет отношения.Для контраста pufA / yakA модель 6 имела наибольшую поддержку среди ненулевых моделей. Эта модель однозначно соответствует нашим интерпретативным моделям. Он утверждает, что yakA является вышестоящим репрессором pufA , и что yakA включается при агрегации. Оба эти заключения согласуются с тем, что было определено ранее относительно ролей, которые эти три гена играют во время развития [22]. YakA репрессирует pufA , который затем перестает репрессировать pkaC .
regA / pkaR был проблематичным, потому что почти все черты поддерживали Модель 8, модель без условий воздействия. Для предыдущих двух случаев мы предположили, что эти черты не лежат ниже по течению пути. Учитывая это предположение, мы могли заключить, что regA и pkaR не участвовали в агрегации. Однако у двух других контрастов было 435 и 528 признаков, поддерживающих Модель 8, а у regA / pkaR – 1497. Из-за этого несоответствия мы предполагаем, что некоторая доля этих генов поддерживает иерархическую модель, соответствующую Модели 8: что один ген является усиливает другой и дезактивируется агрегацией.Согласно ранее опубликованным результатам, regA и pkaR работают вместе, чтобы подавить пре-агрегацию pkaC и фактически деактивированы после агрегации [23]. Это согласуется с потенциальными иерархическими отношениями.
Поскольку мы моделируем неаддитивные взаимодействия, преобразование логарифмического масштаба этих данных может потенциально изменить результаты относительно нетрансформированных данных [3], [24]. Чтобы проверить это, мы возведем в степень данные и повторили наш метод.Несмотря на кардинальные изменения формы распределения данных, полученное распределение наиболее подходящих моделей соответствовало результатам, представленным выше. Опять же, большинство черт не показали никаких эффектов удаления (т.е.поддерживают Модель 8). Модель 2 имела наибольшую поддержку контраста pufA / pkaC , Модель 6 имела наибольшую поддержку контраста pufA / yakA , а Модель 8 почти полностью поддерживала контраст regA / pkaR с использованием данные после агрегирования (рисунок S5).Интересно, что это не означает, что каждая черта поддерживает одну и ту же модель независимо от масштабного преобразования. Фактически, только 57% и 47% признаков поддерживают одну и ту же модель с нетрансформированными данными для контраста pufA / pkaC и pufA / yakA соответственно. Однако в обоих этих случаях подавляющее большинство измененных признаков поддерживают Модель 8. Этот результат изменяет нашу предыдущую интерпретацию признаков, поддерживающих Модель 8; в дополнение к генам, не находящимся ниже по ходу пути, может существовать некоторая доля генов, для которых изменения экспрессии из-за делеции не обнаруживаются из-за проблем с масштабом.Меньшее количество признаков поддерживало Модель 8 с использованием преобразованных данных, что позволяет предположить, что эти данные могут быть более информативными при использовании логарифмического преобразования.
Таким образом, во всех трех случаях наши наилучшие модели регрессии соответствуют набору моделей интерпретации, которые включают истинные отношения между генами. Некоторые регрессионные модели имеют отношение «один ко многим» с интерпретирующими моделями, но в этих случаях количество возможных интерпретирующих моделей сокращается до нескольких. Модель 6 соответствует только одной интерпретации, что позволяет легко описать контраст pufA / yakA .При оценке pufA / pkaC модель 2 соответствует одной иерархической модели и одной модели одного гена. Поскольку контраст pufA / yakA свидетельствует о том, что удаление pufA имеет эффект, иерархическая модель является предпочтительной интерпретацией модели только pkaC . Поскольку мы меняем порог значимости для выбора модели, наши результаты устойчивы. Модель наилучшего соответствия среди моделей 1–7 была одинаковой для пороговых значений p от 0.05 до 0,001 (рисунок S6). По мере того, как критерий отбора становится более строгим, мы отклоняем большее количество эффектов как несущественных, и большее количество признаков поддерживает Модель 8.
Обсуждение
Измерение количества транскриптов в клетке останется фундаментальным интересом для биологов. Технологии экспрессии генов стали популярными за последнее десятилетие из-за их способности захватывать множество генов одновременно. Анализы, которые традиционно фокусировались на нескольких генах, теперь должны быть расширены, чтобы рассмотреть целые геномы.В этом масштабе отношения между генами представляют не меньший интерес, чем индивидуальные эффекты генов. Существует множество методов для вывода генных сетей или путей из профилей экспрессии [25]. Большинство из них требуют больших наборов данных и приводят к большим сетевым диаграммам, которые трудно интерпретировать. Эти подходы полезны, потому что они обеспечивают представление транскрипции в масштабе генома, и они удобны, потому что их можно применять к данным из множества легкодоступных источников.
Тем не менее, существует постоянная потребность в экспериментах, которые позволят нам напрямую вывести пути.Классический эксперимент с эпистазом, который мы описываем в наших результатах [21], является одним из таких подходов. Поскольку он напрямую нацелен на пары генов, мы можем строить пути взаимоотношений за раз. Такой локальный подход приводит к легко понимаемым и биологически релевантным схемам путей. Кроме того, он связывает генетические вариации с вариациями экспрессии. По этим причинам подобные эксперименты будут все более полезными при построении диаграмм биологических систем. Хотя в настоящее время проводится несколько экспериментов, которые измеряют экспрессию в генетически изменчивой популяции, их количество быстро растет.Наша мотивация – предоставить концептуальную основу, в которой можно интерпретировать эти и связанные с ними эксперименты. Мы рассмотрели простейшую структуру генетически изменчивых данных для идентификации эпистаза, при которой индивидуумы различаются только по двум локусам, но наши идеи могут быть применены к ряду аналогичных данных.
Поскольку данные экспрессии по своей природе непрерывны, мы должны обращаться к ним с помощью количественных методов. Регрессионный анализ – это стандартный метод связи непрерывных переменных. Использование модели множественной регрессии для оценки эффектов и взаимодействий генов имеет несколько преимуществ.Во-первых, это позволяет нам рассматривать информацию от всех делеционных мутантов и дикого типа одновременно. Кроме того, он оценивает эффект для каждого аллеля, учитывает дисперсию аллельных эффектов и отделяет эти эффекты от дисперсии ошибок. В традиционном анализе эпистаза двойной мутант сравнивается с каждым одиночным мутантом на основе правил, и два ближайших значения признака определяют эпистаз. В отличие от нашего метода, этот метод не использует всю информацию из заданного контраста, и трудно отличить сигнал от шума.Существует множество сложных методов подгонки множественных регрессионных моделей, и их следует использовать на основе свойств распределения конкретных данных.
Мы рассматриваем индивидуальные особенности экспрессии, а не профиль экспрессии. Модель экспрессии генов представляет каждый признак, но мы должны вывести правильную биологическую модель на основе результатов этапа регрессии. Соответствующая модель регрессии представляет каждую возможную модель экспрессии генов, но эти отношения не всегда однозначны.Иерархии, в которых вышестоящий ген отключается сигналом, смешиваются со случаями, когда ген не имеет никакого эффекта. Имеет смысл, что мы не можем наблюдать эффект делеции, если ген уже выключен у дикого типа. Тем не менее, наша схема во всех случаях соответствовала предыдущим характеристикам пути.
Масштабные преобразования распространены в генетике и геномике, поэтому данные соответствуют допущениям статистического тестирования, таким как нормальность и гомоскадасность [3].Логарифмические преобразования повсеместно встречаются в литературе для данных экспрессии генов, таких как представленные в наших результатах. Однако модели с неаддитивным взаимодействием зависят от масштаба данных, и преобразования могут привести к поддержке альтернативных моделей. Это давняя проблема с описанием эпистаза сложных признаков [24]. Часто бывает трудно узнать наиболее биологически подходящую шкалу, и вместо этого шкала часто выбирается произвольно на основе доступных измерений или статистического удобства.Для признаков экспрессии генов проблема масштаба еще более сложна. Поскольку существуют большие различия в диапазоне вариаций экспрессии между генами, вполне вероятно, что ни одна шкала не позволит выявить лежащие в основе биологические взаимодействия для всех признаков экспрессии. Взаимосвязь между чешуей и эпистазом – это область, которая требует дальнейшего изучения, особенно в эту эпоху генетики, биомолекулярных признаков, таких как экспрессия генов, которые в этом контексте не были хорошо изучены.
Когда мы выполнили такой же анализ логарифмически преобразованных и нетрансформированных данных постагрегации, примерно половина признаков поддерживала другую модель наилучшего соответствия, однако распределение результатов привело к тем же выводам относительно лежащей в основе взаимосвязи между удаленными генами.Это говорит о том, что наши выводы могут быть устойчивыми для масштабирования эффектов, которые могут повлиять на отдельные признаки, поскольку они основаны на распределении всех признаков. Те черты, на которые влияет масштаб, имеют тенденцию к отсутствию обнаруживаемых эффектов удаления с нетрансформированными данными. Это еще больше сбивает с толку примерно одну треть признаков, поддерживающих Модель 8, что также может указывать на восходящий энхансер или признак, на самом деле не затронутый делециями. Хотя мы не обесцениваем эффекты шкалы, мы предполагаем, что большинство этих черт соответствуют последней категории из-за высокого процента этих черт, согласованности черт, поддерживающих Модель 8, между контрастами и логики, согласно которой делеции должны влиять только на последующие гены.В любом случае, эти опасения являются веским аргументом в пользу интерпретации распределения результатов по признакам выражения. Это контрастирует с методами, которые рассматривают все черты как профиль выражения. Они предполагают, что профиль в целом поддерживает один основной путь [21].
Используя наш метод, легко интерпретировать ряд экспериментов. Изучаемые аллели не обязательно должны быть нулевыми аллелями, например удаления. Тот же метод можно применить к сверхэкспрессированным генам или любому полиморфному локусу.Кроме того, в методе можно проводить эксперименты по изучению множественных локусов и взаимодействий более высокого порядка. Трех- и четырехсторонние модели эпистаза следуют тем же принципам, что и двухсторонние модели, которые мы представляем. Модель регрессии очень гибкая и ее легко расширить, добавив параметр для каждого локуса плюс условия взаимодействия. Связь этих статистических моделей с биологическими моделями происходит по тому же процессу, который мы проиллюстрировали. Сильные стороны нашего подхода особенно очевидны в моделях с несколькими локусами, потому что мы предоставляем средства для оценки эффектов с использованием всей популяции мутантов одновременно.Количество генотипов увеличивается в два раза для каждого дополнительного гена, включенного в эксперимент; с экспериментом с тремя локусами, имеющим восемь генотипов. По мере увеличения количества необходимых парных сравнений они будут содержать больше необнаруженных ошибок и их будет труднее интерпретировать. Воздействие окружающей среды также может быть включено в модель за счет увеличения сложности интерпретации. Мы рассмотрели наблюдения до агрегирования и после агрегирования отдельно в нашем примере для простоты.
Продолжая добавлять генетическую и экологическую сложность, становится очевидным, как классическая структура эпистаза связана с количественной генетической парадигмой. Дополнительным преимуществом нашего метода является то, что он позволяет проводить сравнения между любыми популяционными анализами экспрессии. Независимо от того, состоят ли исследуемые популяции из делеционных мутантов, экспериментально разработанных скрещиваний, инбредных линий, штаммов с заменой хромосом или природных популяций, все признаки экспрессии одинаковы. По этой причине очень желательно сравнение этих результатов.Оценка аллельных эффектов и взаимодействий для каждого признака экспрессии позволяет напрямую сравнивать различные генетические фоны. Приняв общую интерпретирующую структуру для ряда экспериментов, в которых экспрессия генов используется в качестве признака, мы можем интегрировать результаты и сформировать более четкое представление о генетическом контроле систем.
Материалы и методы
Данные экспрессии гена Dictyostellium
Мы использовали данные, первоначально представленные Van Driessche et al. Мы используем данные из Dictyostellium discoideum дикого типа [20] и восьми штаммов мутантных штаммов с делециями ( pufA –, pkaC – , pufA – pkaC – ,
6–
pufA – yakA – , regA – , pkaR – , regA – pkaR – ) [21].Они измерили профиль экспрессии генов каждого штамма с течением времени, используя микрочипы кДНК и общий эталон, который был объединен для всех временных точек. Экспрессию измеряли тринадцать раз в течение 24 часов и регистрировали реакцию агрегации развития на лишение питательных веществ, которая начиналась в середине временного цикла. Мы сгруппировали наблюдения до (0,2,4,6 часа) и после (14,16,18,20 часов) агрегирования. Экспрессия в эти моменты времени сильно коррелирована ([Рисунок 2 в 20]) и согласуется с регуляторными изменениями, о которых сообщалось ранее.Этот пул данных увеличил размер выборки для нашего регрессионного анализа. Наблюдения во время переходного периода (часы 8, 10 и 12) не принимались во внимание, как и наблюдения на поздних стадиях развития, которые были менее коррелированными (часы 22 и 24). Данные представляют кратное изменение в логарифмической шкале. Мы изучили 1553 гена, которые были измерены в обоих экспериментах и дифференциально экспрессировались в организме дикого типа во время агрегации [20].Регрессионный анализ
Мы подбираем модели в статистической среде R [26].Пошаговый выбор в обратном порядке влечет за собой подгонку полностью параметризованной модели с последующим удалением членов модели, которые не соответствуют заданному порогу значимости. Модель модифицируется с использованием оставшихся условий до тех пор, пока не будут исключены никакие другие условия.
Благодарности
Спасибо Эрику Стоуну и Игнацио Карбоне за полезные обсуждения и комментарии к нашей рукописи. Спасибо также двум анонимным рецензентам за их комментарии и предложения.
Вклад авторов
Задумал и спроектировал эксперименты: DA ZZ.Проанализированы данные: DA. Написал бумагу: Д.А.
Ссылки
- 1. Филлипс П.С. (1998) Язык взаимодействия генов. Генетика 149: 1167–1171.
- 2. Хьюз Т.Р., Мартон М.Дж., Джонс А.Р., Робертс С.Дж., Стоутон Р. и др. (2000) Функциональное открытие через сборник профилей экспрессии. Cell 102: 109–126.
- 3. Линч М., Уолш Б. (1998) Генетика и анализ количественных признаков. Сандерленд, Массачусетс: Sinauer Associates.
- 4. Тонг А.Х., Лесаж Дж., Бадер Г.Д., Динг Х., Сюй Х. и др. (2004) Глобальное картирование сети генетического взаимодействия дрожжей. Наука 303: 808–813.
- 5. Тонг А.Х., Евангелиста М., Парсонс А.Б., Сюй Х., Бадер Г.Д. и др. (2001) Систематический генетический анализ с упорядоченными массивами делеционных мутантов дрожжей. Наука 294: 2364–2368.
- 6. St Onge RP, Mani R, Oh J, Proctor M, Fung E и др. (2007) Систематический анализ путей с использованием профилей пригодности комбинаторных делеций генов с высоким разрешением.Нат Генет 39: 199–206.
- 7. Jansen RC, Nap JP (2001) Генетическая геномика: добавленная стоимость от сегрегации. Тенденции Genet 17: 388–391.
- 8. Schadt EE, Monks SA, Drake TA, Lusis AJ, Che N, et al. (2003) Генетика экспрессии генов, исследованная у кукурузы, мышей и человека. Природа 422: 297–302.
- 9. Brem RB, Yvert G, Clinton R, Kruglyak L (2002) Генетическое вскрытие регуляции транскрипции у почкующихся дрожжей. Наука 296: 752–755.
- 10. Быстрых Л., Веерсинг Э., Донтье Б., Саттон С., Плетчер М.Т. и др. (2005) Выявление регуляторных путей, которые влияют на функцию гемопоэтических стволовых клеток, с использованием «генетической геномики». Нат Генет 37: 225–232.
- 11. Chesler EJ, Lu L, Shou S, Qu Y, Gu J и др. (2005) Комплексный анализ признаков экспрессии генов раскрывает полигенные и плейотропные сети, которые модулируют функцию нервной системы. Нат Генет 37: 233–242.
- 12. Ли Дж., Бурмейстер М. (2005) Генетическая геномика: сочетание генетики с анализом экспрессии генов.Hum Mol Genet 14 Спец. № 2: R163–169.
- 13. Storey JD, Akey JM, Kruglyak L (2005) Анализ множественных локусных сцеплений экспрессии в масштабе всего генома в дрожжах. PLoS Biol 3: e267.
- 14. Auger DL, Gray AD, Ream TS, Kato A, Coe EH Jr и др. (2005) Неаддитивная экспрессия генов у диплоидных и триплоидных гибридов кукурузы. Генетика 169: 389–397.
- 15. Гибсон Дж., Райли-Бергер Р., Харшман Л., Копп А., Вача С. и др. (2004) Обширная неаддитивность экспрессии генов в зависимости от пола у Drosophila melanogaster.Генетика 167: 1791–1799.
- 16. Гибсон Г., Вейр Б. (2005) Количественная генетика транскрипции. Тенденции Genet 21: 616–623.
- 17. Брем Р. Б., Кругляк Л. (2005) Пейзаж генетической сложности 5700 признаков экспрессии генов у дрожжей. Proc Natl Acad Sci U S A 102: 1572–1577.
- 18. Эйвери Л., Вассерман С. (1992) Упорядочивающая функция гена: интерпретация эпистаза в регуляторных иерархиях. Тенденции Genet 8: 312–316.
- 19. Ли Р., Цай С.В., Шокли К., Стилиану И.М., Вергедал Дж. И др. (2006) Структурный модельный анализ множества количественных характеристик. PLoS Genet 2: e114.
- 20. Ван Дрише Н., Шоу С., Като М., Морио Т., Сукганг Р. и др. (2002) Транскрипционный профиль многоклеточного развития у Dictyostelium discoideum. Развитие 129: 1543–1552.
- 21. Ван Дрише Н., Демсар Дж., Бут Е.О., Хилл П., Джуван П. и др. (2005) Анализ эпистаза с глобальными транскрипционными фенотипами.Нат Генет 37: 471–477.
- 22. Souza GM, da Silva AM, Kuspa A (1999) Голодание способствует развитию Dictyostelium, снимая ингибирование PufA трансляции PKA через путь киназы YakA. Разработка 126: 3263–3274.
- 23. Shaulsky G, Fuller D, Loomis WF (1998) цАМФ-фосфодиэстераза контролирует PKA-зависимую дифференцировку. Развитие 125: 691–699.
- 24. Франкель В. Н., Шорк Н. Дж. (1996) Кто боится эпистаза? Нат Генет 14: 371–373.
- 25. Bansal M, Belcastro V, Ambesi-Impiombato A, di Bernardo D (2007) Как вывести генные сети из профилей экспрессии. Мол Сист Биол 3: 78
- 26. Основная группа разработчиков R (2007) R: Язык и среда для статистических вычислений. Вена, Австрия: Фонд R для статистических вычислений. ISBN 3-1-07-0.
ESP Основы классической генетики
Серия основ ESP 29 октября 2021 г. Обновлено:
Классическая генетика
Проект электронных научных публикаций был первоначально создан для обеспечить доступ к важным работам, связанным с основами классическая генетика.Общий сайт WWW.ESP.ORG теперь расширен. для обеспечения доступа и к другим материалам. Эта текущая страница имеет теперь стать корневой страницей раздела основ классической генетики.
Следите за новыми публикациями, изменениями на сайте и другими новостями.
Просмотрите нашу аннотированную коллекцию статей и книг, связанных с генетика.Полнотекстовые PDF-версии работ из этого сборника представлены доступно на сайте ESP.
Просмотрите нашу аннотированную коллекцию полных цифровых книг по теме генетике и другим областям.
Просмотрите нашу коллекцию биографических материалов об ученых занимается классической генетикой и другими областями, представляющими интерес для Научно-издательский проект «Электроника».
Генетика в контексте предлагает двойную хронологию с научными событиями (особенно те, которые имеют отношение к классической генетике), представленные против представляющие общий исторический и культурный интерес. Динамические ссылки предоставлены соответствующие документы на сайте ESP и в других местах.
Эта функция дает исторический контекст для вех в истории генетики. Например, знаете ли вы, что Дарвин опубликовал Происхождение видов за год до избрания Линкольна Президент, а Мендель опубликовал свои выводы через год после Линкольн был убит?
Ознакомьтесь с этими и другими событиями в Genetics in Context
История и цели проекта ESP
Вступление
Хотя проблемы наследования интересовали наблюдателей с тех пор, как начало письменной истории, систематический, научный изучение наследственности началось только в конце девятнадцатого века, затем резко вырос как интеллектуальное поле вскоре после 1900 года.В трансформация генетики из наблюдательных, анекдотических попыток к истинной науке со строгими, глубокими теоретическими основами и с глубокой объяснительной силой произошло очень быстро.
Классическая генетика, изучение закономерностей наследования, началась с повторное открытие работ Менделя в 1900 году и процветало в первая половина века.
Потому что генетика развивалась так быстро всего за несколько десятилетий после 1900 г., литература того периода представляет собой ценный ресурс. даже сейчас.Его могут с пользой прочитать студенты и ученые, желающие чтобы понять основы своей области, а также непрофессионалами или историки науки.
К сожалению, раннюю литературу становится все труднее найти. В нынешних учебниках едва ли посвящена вводная глава классической генетики, и эти презентации существенно мифологизировал ранние работы, так что настоящее понимание потеряно.Было разработано несколько сборников классических произведений годы (хотя и не в последнее время), но они страдают от последствий необходимых, но, тем не менее, вредных, очень избирательных выборка, которая сопровождает эти проекты. Такая селективность в сочетании с введениями, которые предлагают по существу современные интерпретации работы, затемняют интеллектуальную строгость и азарт оригинальные усилия.
Чтобы решить эти проблемы, The Electronic Scholoarly Publishing Проект был создан для переиздания классической литературы по генетике в электронная форма, так что доступ к этим важным работам будет тривиально просто для всех, у кого есть доступ к простому просмотру веб-страниц программное обеспечение.
Литература по классической генетике остается интеллектуально доступной. всем читателям, отчасти потому, что все читатели приносят с собой готовые знакомство с фактом наследования (вроде как порождает, еще или меньше) и нужно только понять механизмы наследственность как биологический процесс.Хотя современная молекулярная биология включает в себя методы, которые непосвященным могут показаться почти волшебными, оригинальные методики классической генетики по достоинству оценят все как один: скрещивание особей, различающихся каким-либо наследственным признаком, собрать всех потомков, оценить их атрибуты и предложить механизмы, объясняющие наблюдаемые закономерности наследования. Повторить как необходимо.
Читая ранние работы классической генетики, можно почти неумолимо, во все более сложные модели, пока молекулярные объяснения начинают казаться необходимыми и естественными.В таком случае, инструменты для понимания современных молекулярных исследований уже под рукой.
Цель этого проекта – помочь читателям достичь этой точки.
В начале 1990-х гг. Роберт Роббинс был преподавателем в Johns Хопкинса, где он руководил информационным ядром GDB. – база данных картирования генов человека международного геномный проект.Чтобы поделиться документами с коллегами по всему миру, он создал на своей личной веб-странице небольшой раздел для обмена бумагами. Этот небольшой проект превратился в Electronic Scholarly Издательский проект.
В 1995 году Роббинс стал вице-президентом по информационным технологиям отдела исследований рака Фреда Хатчинсона. Центр в Сиэтле, штат Вашингтон. Вскоре после прибытия в Сиэтл Роббинс обеспечил финансирование через компонент ELSI Проекта генома человека в США для создать оригинальный ESP.Веб-сайт ORG с формальной целью предоставление бесплатного всемирного доступа к литературе по классической генетике.
Хотя методы молекулярной биологии могут показаться почти волшебный для непосвященных, оригинал методы классической генетики по достоинству оценят все: скрестить особей, различающихся каким-либо наследственным признаком, собрать всех потомков, оцените их атрибуты и предложите механизмы чтобы объяснить наблюдаемые закономерности наследования.
Читая ранние работы классической генетики, можно почти неумолимо, во все более сложные модели, пока молекулярные объяснения начинают казаться и нужным, и естественным. На этом этапе инструменты для понимания генома исследования под рукой. Помощь читателям в достижении этот момент был первоначальной целью The Electronic Scholarly Publishing. Проект.
Использование сайта быстро росло и остается высоким. Факультет начал использовать сайт для назначенных им чтений. Другое в сети издатели, от The New York Times до Nature ссылались на материалы ESP в своих публикациях. Нобелевские лауреаты (например, Джошуа Ледерберг) регулярно использовал сайт и даже написал, чтобы предложить изменения и улучшения.
Когда сайт начинался, журналов не было. делали свой ранний контент доступным в цифровой формат. В результате ESP была вынуждена оцифровать классический литературу до того, как она стала доступной. Для многих важных документы – такие как Оригинальная статья Менделя или первая генетическая карта – ESP пришлось выпускать совершенно новые наборные версии произведений, если бы они были доступны в высококачественном формате.
Ранняя поддержка со стороны Министерства энергетики проекта «Геном человека» была критически важно для получения прочного фундамента проекта ESP. С тех пор, как это финансирование закончилось (почти 20 лет назад), проект был действовали исключительно на добровольных началах. Всем, кто желает помочь в этих усилиях, следует прислать электронное письмо Роббинсу.
(PDF) История классической генетики
История классической генетики
синоним, тогда что же делает фактор, чтобы выявить
о признаке? Морган и его Fly Group, а также в значительной степени
большая часть американских генетиков приняли сознательное решение сосредоточиться, по крайней мере экспериментально, если не мысленно, на генетике передачи
(как гены передаются от поколения к поколению). ) на
за счет развития (как гены играют роль в влиянии
на рост отдельного организма).Недавние исторические исследования показали, что это повествование имеет тенденцию исключать гораздо более широкую концепцию генетики, практикуемую, в частности, в Германии, в частности, в Германии, где физиологическая генетика и генетика развития оставались
центральными исследованиями на протяжении первой половины периода. ХХ
век. Более того, члены кружка Моргана использовали мощные инструменты, которые они разработали для изучения генетики плодов, для решения
вопросов развития, эволюции и других биологических вопросов, выходящих за рамки законов передачи.Классическая генетика середины века,
, однако, часто воспринимается как наука о передаче генетики, занимающаяся прежде всего объяснением закономерностей наследования.
Первые важные сведения о действии генов появились в Англии и из неожиданного для медицины сектора
. Вылечив
пациентов, страдающих алкаптонурией, заболеванием, которое в дальнейшем вызывает болезненный артрит
, врач Арчибальд Гаррод показал, что
заболевание следует по менделевскому типу наследования, а
проявляется в отсутствии фермента, специализирующегося на ферментах. в разложении гомогентистической кислоты
(которая выделяется с мочой
пациентов с алкаптонурией и становится черной при воздействии воздуха
, что является характерным признаком слияния с заболеванием).Поскольку паттерн
наследования заболевания предполагал, что действует единственный рецессивный ген
, Гаррод пришел к выводу, что неспособность синтезировать фермент
должна быть вызвана дефектом в гене. В лекции
Королевского колледжа врачей в 1908 году Гаррод предположил, что алкаптонурия
является «врожденной ошибкой метаболизма», и придумал поговорку
«один ген, один фермент», чтобы описать, что на самом деле делают гены.
См. Также: Гены: определение и структура.
Neurospora crassa.Работая с российским генетиком
Борисом Эфрусси в Париже в 1930-х годах над генетическим определением цвета глаз у дрозофилы, Бидл убедился, что гены
каким-то образом формируют биохимические пути. Он и Татум теперь
облучили Neurospora рентгеновскими лучами и показали, что плесневые грибки с
определенными дефектными генами из-за радиации не могут производить ферменты
, необходимые для расщепления простых элементов
их рациона.Гаррод изучал естественный генетический дефект
у людей, тогда как Бидл и Татум изучали искусственно индуцированный
дефект у гриба, но оба они достигли одного и того же скопления
: один ген, один фермент. Этот результат имел тенденцию поддерживать выводы
, сделанные J.S. Хаксли в своей более ранней работе продемонстрировал действие «генов скорости», влияющих на цвет глаз у пресноводных креветок,
, Gammarus, а Р. Скотт-Монскрифф основала
на своем исследовании генетики Цветная пигментация у различных видов
штаммов львиного зева, Antirrhinums.В 1950-х годах американский химик Линус Полинг (Linus Pauling) обнаружил, что серповидно-клеточная анемия у людей
вызвана дефектом гемоглобина и что болезнь
наследуется простым, менделевским способом. Таким образом, Полинг обобщил
пословицу Гаррода еще дальше, и «один ген, один фермент»
теперь стал одним геном, одним белком. См. Также: Бидл, Джордж
Уэллс; Полинг, Линус Карл; Татум, Эдвард Лори
Из чего состоят гены?
Наряду с исследованием того, что на самом деле делают гены, было проведено исследование
того, из чего они сделаны.Морган постулировал
как физическое существование генов в хромосомах, но это
было основано на статистическом выводе, полученном из результатов
экспериментов по разведению. Первой вехой на пути к выяснению физической природы гена
стал бывший ученик
Моргана Герман Мюллер, который в 1927 году показал
, что гены являются реальными физическими сущностями, открыв хромосомные
некоторые. детей на рентгеновские лучи и наблюдая, что физические изменения хромосом
, вызванные излучением, привели к мутациям
, выраженным в фенотипе детей.Макс Дельбрюк, физик
, ставший генетиком, работавший в Берлине в середине 1930-х годов, объединил
рентгеновские экспериментальные данные и квантово-механические рассуждения, чтобы объяснить
мутации с точки зрения химического и физического поведения атомов. Дельбрюк переехал в Соединенные Штаты в 1937 г.
и начал независимое сотрудничество с итальянским беженцем
, генетиком Сальвадором Луриа и химиком Альфредом Херши, в результате чего
привело к образованию Phage Group, неофициальной сети, посвященной вирусным и вирусным исследованиям. бактериальные генетики сосредоточились на изучении бактериофага
– крошечного вируса, который заражает бактерии и быстро размножается
внутри хозяина.Физик Дельбрюк представил эту систему
как биологический аналог атома водорода: sim-
, простой для изучения и содержащий информацию о природе
передачи и ее материале. См. Также: Cold Spring Har-
bor Laboratory; Delbr ̈
uck, Max Ludwig Henning; Hershey,
Alfred Day; History of Bacteriology; History of Molecular
Biology; Muller, Hermann Joseph; Mutagenesis
What genes из чего были сделаны до сих пор неизвестно.Работа Бидла
и Татума, поскольку она связывала гены с ферментами,
имела тенденцию укреплять широко распространенное мнение о том, что гены были сделаны из белка
. Если бы они были сконцентрированы в ядре, гены
могли бы быть либо белком, либо ДНК, а химический анализ показал, что
ДНК была намного более простой молекулой, чем белок, следовательно, менее
кандидат на носитель генетической информации. Но в 1944 году
группа в Институте медицинских исследований Рокфеллера во главе с
Освальдом Т.Компания Avery сообщила, что ей удалось трансформировать
a неинфекционных бактерий в инфекционные, она тщательно выделила
«трансформирующий» фактор, и этим фактором была ДНК.
пожилой джентльмен с уходом на пенсию, Эйвери уклонялся от
настаивать на сути, и большинство биологов продолжали верить
, что гены состоят из белка. Однако в 1952 году Херши и
Марта Чейз, оба члены группы фагов, показали, что
, когда бактериофаг заражает бактериальную клетку, только ДНК и ДНК
вводятся в бактерии; белковая оболочка вируса
неизменно остается снаружи, выбрасывается.Это было убедительным доказательством того, что ДНК на самом деле была генетическим материалом. См. Также: Avery,
Oswald Theodore; Macleod, John James Richard; McCarty,
Maclyn; Nucleic Acids as Genetic Material
.В этот период метафора информации приобрела популярность в биологии.