
%d0%ba%d0%be%d0%bd%d1%82%d1%80%d0%be%d0%bb%d1%8c%d0%bd%d0%b0%d1%8f%20%d1%80%d0%b0%d0%b1%d0%be%d1%82%d0%b0 — со всех языков на все языки
Все языкиАбхазскийАдыгейскийАфрикаансАйнский языкАканАлтайскийАрагонскийАрабскийАстурийскийАймараАзербайджанскийБашкирскийБагобоБелорусскийБолгарскийТибетскийБурятскийКаталанскийЧеченскийШорскийЧерокиШайенскогоКриЧешскийКрымскотатарскийЦерковнославянский (Старославянский)ЧувашскийВаллийскийДатскийНемецкийДолганскийГреческийАнглийскийЭсперантоИспанскийЭстонскийБаскскийЭвенкийскийПерсидскийФинскийФарерскийФранцузскийИрландскийГэльскийГуараниКлингонскийЭльзасскийИвритХиндиХорватскийВерхнелужицкийГаитянскийВенгерскийАрмянскийИндонезийскийИнупиакИнгушскийИсландскийИтальянскийЯпонскийГрузинскийКарачаевскийЧеркесскийКазахскийКхмерскийКорейскийКумыкскийКурдскийКомиКиргизскийЛатинскийЛюксембургскийСефардскийЛингалаЛитовскийЛатышскийМаньчжурскийМикенскийМокшанскийМаориМарийскийМакедонскийКомиМонгольскийМалайскийМайяЭрзянскийНидерландскийНорвежскийНауатльОрокскийНогайскийОсетинскийОсманскийПенджабскийПалиПольскийПапьяментоДревнерусский языкПортугальскийКечуаКвеньяРумынский, МолдавскийАрумынскийРусскийСанскритСеверносаамскийЯкутскийСловацкийСловенскийАлбанскийСербскийШведскийСуахилиШумерскийСилезскийТофаларскийТаджикскийТайскийТуркменскийТагальскийТурецкийТатарскийТувинскийТвиУдмурдскийУйгурскийУкраинскийУрдуУрумскийУзбекскийВьетнамскийВепсскийВарайскийЮпийскийИдишЙорубаКитайский
Все языкиАбхазскийАдыгейскийАфрикаансАйнский языкАлтайскийАрабскийАварскийАймараАзербайджанскийБашкирскийБелорусскийБолгарскийКаталанскийЧеченскийЧаморроШорскийЧерокиЧешскийКрымскотатарскийЦерковнославянский (Старославянский)ЧувашскийДатскийНемецкийГреческийАнглийскийЭсперантоИспанскийЭстонскийБаскскийЭвенкийскийПерсидскийФинскийФарерскийФранцузскийИрландскийГалисийскийКлингонскийЭльзасскийИвритХиндиХорватскийГаитянскийВенгерскийАрмянскийИндонезийскийИнгушскийИсландскийИтальянскийИжорскийЯпонскийЛожбанГрузинскийКарачаевскийКазахскийКхмерскийКорейскийКумыкскийКурдскийЛатинскийЛингалаЛитовскийЛатышскийМокшанскийМаориМарийскийМакедонскийМонгольскийМалайскийМальтийскийМайяЭрзянскийНидерландскийНорвежскийОсетинскийПенджабскийПалиПольскийПапьяментоДревнерусский языкПуштуПортугальскийКечуаКвеньяРумынский, МолдавскийРусскийЯкутскийСловацкийСловенскийАлбанскийСербскийШведскийСуахилиТамильскийТаджикскийТайскийТуркменскийТагальскийТурецкийТатарскийУдмурдскийУйгурскийУкраинскийУрдуУрумскийУзбекскийВодскийВьетнамскийВепсскийИдишЙорубаКитайский
Итоговая аттестация | Реализация Федерального закона «Об образовании в Российской Федерации»
Введенные Министерством образования и науки РФ Всероссийские проверочные работы (далее – ВПР) имеют своей целью прежде всего обеспечение единства образовательного пространства на территории РФ путем предоставления образовательным организациям единых критериев оценивания качества усвоения обучающимися учебных дисциплин, основанных на Федеральных государственных образовательных стандартах (ФГОС).
В соответствии с утвержденным 15 декабря 2016 года Министерством образования и науки РФ Комплексом мер, направленных на систематическое обновление содержания общего образования на основе результатов мониторинговых исследований и с учетом современных достижений науки и технологий, изменений запросов учащихся и общества, ориентированности на применение знаний, умений и навыков в реальных жизненных условиях, предусмотрено проведение мониторинговых исследований качества общего образования.
В соответствии с Приказом Министерства образования и науки РФ «О проведении мониторинга качества общего образования» от 20 октября 2017 года № 1025, в 2017/2018 учебном году предусмотрено проведение мониторинга качества подготовки обучающихся 4-11 классов по ряду дисциплин в форме проверочных работ.
Информационное письмо Федеральной службы по надзору в сфере образования и науки (Рособрнадзора) от 17 января 2017 года № 05-11 разъясняет, что:
– участие образовательных организаций в проведении мониторинга в форме ВПР является обязательным для 4-5 классов, но не является таковым для проведения мониторинга в 6-11 классах;
– ВПР для выпускников 11-х классов проводится по тем учебным предметам, которые они не выбрали тот или иной предмет для прохождения государственной итоговой аттестации в форме единого государственного экзамена;
– ВПР проводится образовательной организацией самостоятельно на основе единых вариантов заданий, разрабатываемых Федеральным институтом педагогических измерений, которые должны дать возможность оценить результаты обучающихся по единым критериям;
– Рособрнадзор не рекомендует образовательным организациям использовать результаты ВПР для выставления годовых отметок обучающимся;
– результаты ВПР могут быть полезны родителям и педагогам для оценки текущего уровня подготовки обучающихся;
– решение об участии в ВПР конкретного обучающегося принимает образовательная организация.
Отметим, что мнение Рособрнадзора в данной ситуации представляется однозначным и не допускает иных трактовок. Результат выполнения ВПР является независимой оценкой уровня подготовки каждого обучающегося, имеющей цель получения объективных сведений о качестве преподавания и об уровне усвоения обучающимися учебных предметов в образовательной организации.
В соответствии со статьей 58 Закона «Об образовании в Российской Федерации» № 273-ФЗ, промежуточная аттестация проводится в формах, определенных в локальном акте образовательной организации, и в порядке, установленном образовательной организацией. При этом форма и порядок проведения ВПР (в том числе и дата проведения), как уже было сказано, устанавливаются непосредственно Рособрнадзором и Приказами Министерства образования и науки РФ. Со своей стороны, образовательная организация вправе либо согласиться, либо отказаться от проведения ВПР для учащихся 6-11 классов.
Если образовательная организация соглашается провести проверочную работу, то на ее усмотрение итоговые контрольные работы, проводящиеся учителями-предметниками в соответствии со своей рабочей программой, действительно могут быть заменены ВПР (для исключения чрезмерной нагрузки обучающихся).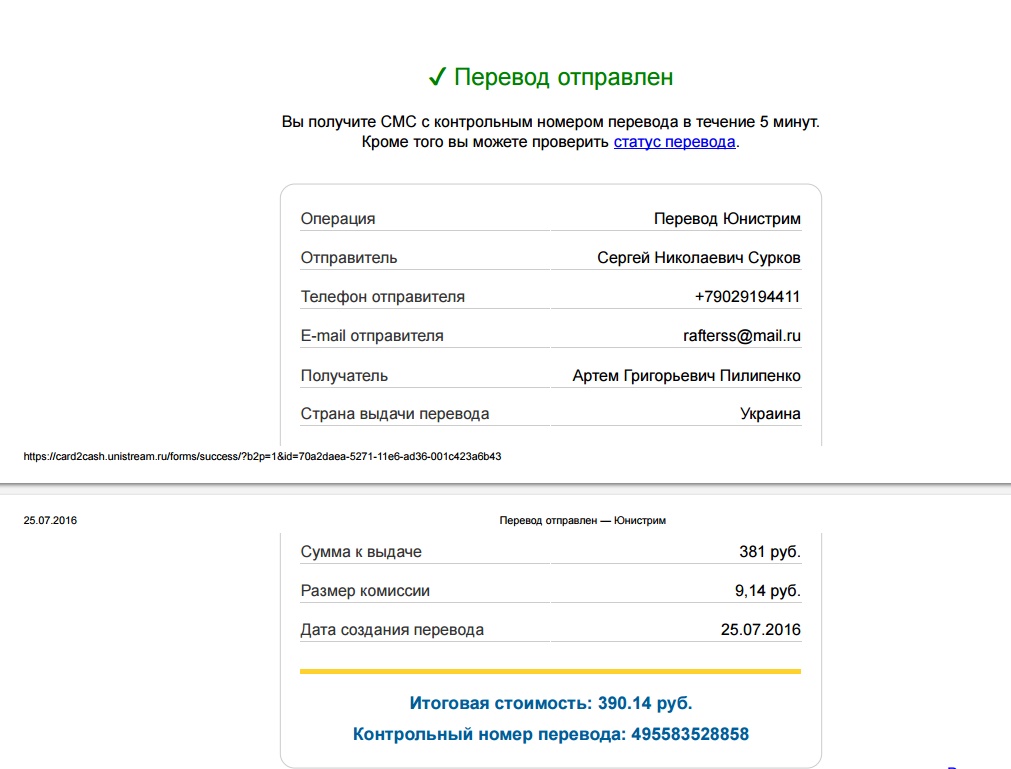
Если же проведение ВПР обязательно (для 4-5 классов), результаты выполнения данной работы могут являться лишь составляющей четвертной и годовой отметки обучающегося (наравне с четвертными и годовой контрольными работами). В данном случае рекомендация Рособрнадзора соблюдается, так как ВПР влияет на годовую отметку не напрямую, а лишь как составляющая прочих форм контроля. Не следует забывать и о том, что в компетенции образовательной организации находится в том числе и внесение полученной обучающимся оценки за ВПР в классный журнал.
Таким образом, ВПР является лишь средством сбора объективных статистических данных об успеваемости обучающихся, но не формой контроля как таковой. Образовательная организация вправе учитывать результат ВПР наравне с текущими контрольными работами, проводимыми ею самостоятельно, но не вправе замещать промежуточную аттестацию.
Кроме того, следует заметить, что непрохождение промежуточной аттестации может привести к условному переводу обучающегося в следующий класс, так как согласно п. 8 статьи 58 Закона «Об образовании в Российской Федерации» № 273-ФЗ обучающиеся, не прошедшие промежуточную аттестацию, переводятся в следующий класс условно, с получением академической задолженности, а согласно пунктам 3 обучающиеся обязаны ликвидировать академическую задолженность.
Ответ подготовлен при участии студента 4 курса факультета права НИУ ВШЭ: Новоселова Ивана Сергеевича
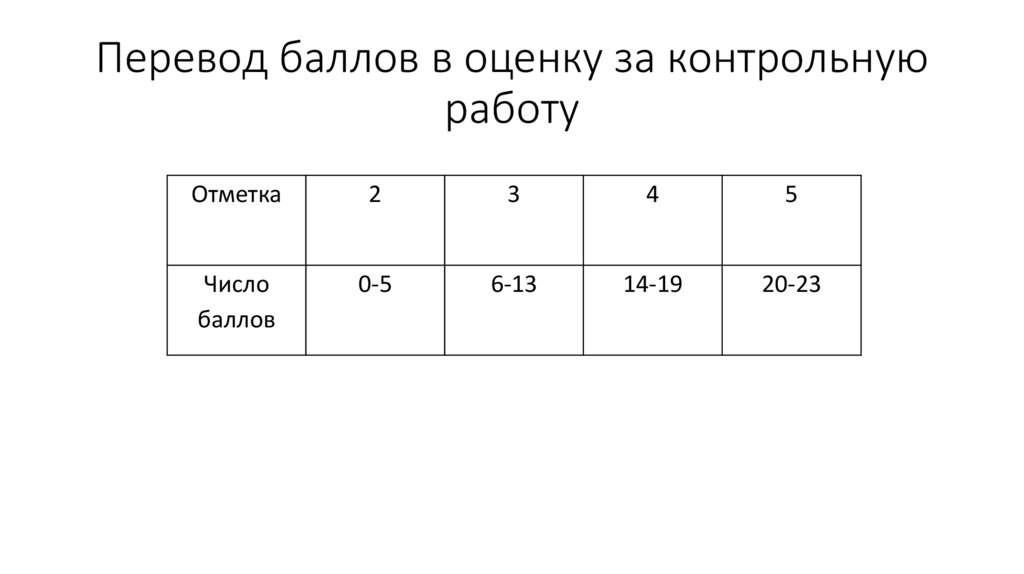
Контрольная работа по теме «Перевод чисел из одной системы в другую. Двоичная арифметика».

Контрольная работа по теме «Перевод чисел из одной системы в другую. Двоичная арифметика». 1 вариант
| Контрольная работа по теме «Перевод чисел из одной системы в другую. Двоичная арифметика». 3 вариант
|
Контрольная работа по теме «Перевод чисел из одной системы в другую. Двоичная арифметика». 2 вариант
. | Контрольная работа по теме «Перевод чисел из одной системы в другую. 4 вариант
. |

 Двоичная арифметика».
Двоичная арифметика».«Перевод чисел из одной системы в другую. Двоичная арифметика».
1 вариант
Перевести десятичное число 9910 в двоичную, восьмеричную и шестнадцатиричную системы счисления.
Перевести двоичное число 10102 в десятичную систему счисления.
Сложить двоичные числа 101112 и 010102.
Из двоичного числа 100112 вычесть число 110002.
Контрольная работа по теме
«Перевод чисел из одной системы в другую. Двоичная арифметика».
3 вариант
Перевести десятичное число 12310 в двоичную, восьмеричную и шестнадцатиричную системы счисления.
Перевести двоичное число 11102 в десятичную систему счисления.
Сложить двоичные числа 011102 и 000112.
Из двоичного числа 111012 вычесть число 100112.
Контрольная работа по теме
«Перевод чисел из одной системы в другую. Двоичная арифметика».
2 вариант
Перевести десятичное число 10110 в двоичную, восьмеричную и шестнадцатиричную системы счисления.

Перевести двоичное число 01012 в десятичную систему счисления.
Сложить двоичные числа 100012 и 101012.
Из двоичного числа 110012 вычесть число 000112
.
Контрольная работа по теме
«Перевод чисел из одной системы в другую. Двоичная арифметика».
4 вариант
Перевести десятичное число 10710 в двоичную, восьмеричную и шестнадцатиричную системы счисления.
Перевести двоичное число 11112 в десятичную систему счисления.
Сложить двоичные числа 111112 и 010112.
Из двоичного числа 101002 вычесть число 101012
.
Контрольная работа по теме «Перевод чисел из одной системы в другую. Двоичная арифметика».
Двоичная арифметика».
1 вариант
Перевести десятичное число 9910 в двоичную, восьмеричную и шестнадцатиричную системы счисления.
Перевести двоичное число 10102 в десятичную систему счисления.
Сложить двоичные числа 101112 и 010102.
Из двоичного числа 100112 вычесть число 110002.
Контрольная работа по теме
«Перевод чисел из одной системы в другую. Двоичная арифметика».
3 вариант
Перевести десятичное число 12310 в двоичную, восьмеричную и шестнадцатиричную системы счисления.
Перевести двоичное число 11102 в десятичную систему счисления.
Сложить двоичные числа 011102 и 000112.

Из двоичного числа 111012 вычесть число 100112.
Контрольная работа по теме
«Перевод чисел из одной системы в другую. Двоичная арифметика».
2 вариант
Перевести десятичное число 10110 в двоичную, восьмеричную и шестнадцатиричную системы счисления.
Перевести двоичное число 01012 в десятичную систему счисления.
Сложить двоичные числа 100012 и 101012.
Из двоичного числа 110012 вычесть число 000112
.
Контрольная работа по теме
«Перевод чисел из одной системы в другую. Двоичная арифметика».
4 вариант
Перевести десятичное число 10710 в двоичную, восьмеричную и шестнадцатиричную системы счисления.

Перевести двоичное число 11112 в десятичную систему счисления.
Сложить двоичные числа 111112 и 010112.
Из двоичного числа 101002 вычесть число 101012
.
Дополнительные меры для борьбы с СOVID-19 вводятся в Свердловской области
02.11.2021г.
В Свердловской области вводятся дополнительные меры для противодействия распространению коронавирусной инфекции. Сложившаяся эпидситуация требует усиления установленного в регионе режима. Такое решение принято 2 ноября.
Кроме того, если контрольные группы в течение следующих трёх дней продолжат фиксировать нарушения на предприятиях торговли, общественного питания и в других зонах, свободных от COVID-19, меры могут быть ужесточены – вплоть до ввода локдауна.
Отметим, контрольные группы в Свердловской области сейчас работают с дополнительной нагрузкой. Всего ежедневно функционируют в регионе 362 контрольные группы с общим количеством участников в 900 человек. На текущий момент проверено 4,5 тысячи объектов потребительского рынка, торговых и торгово-развлекательных центров. Также проверяются общественный транспорт, объекты культуры, физкультуры и спорта.
Всего ежедневно функционируют в регионе 362 контрольные группы с общим количеством участников в 900 человек. На текущий момент проверено 4,5 тысячи объектов потребительского рынка, торговых и торгово-развлекательных центров. Также проверяются общественный транспорт, объекты культуры, физкультуры и спорта.
К проверкам подключаются сотрудники полиции и Росгвардии. Через три дня им предстоит обобщить собранный материал и сделать выводы о результатах инспекции для определения дальнейших шагов. Тем не менее, проверочные мероприятия будут продолжаться и после.
В настоящий же момент прорабатываются следующие изменения в указ №100 о режиме повышенной готовности.
Во-первых, региональные власти рекомендуют работодателям перевести всех не привитых первым компонентом сотрудников на дистант. Будет установлен запрет на вход детей до 18 лет без сопровождения родителей или законных представителей в торгово-развлекательные центры. Эта мера позволит в праздники, например, избежать скопления подростков в закрытых помещениях, поскольку в ТРЦ попадут только те дети, которые идут вместе с родителями за покупками, в кинотеатр или на другие COVID-19-free площадки. Срок, по которому люди могут вместо QR-кода предъявлять справку о вакцинации первым компонентом, предлагается сдвинуть с 15 декабря на 1 декабря. Отдельным решением станет перевод учреждений среднего профессионального образования на дистанционное обучение.
Срок, по которому люди могут вместо QR-кода предъявлять справку о вакцинации первым компонентом, предлагается сдвинуть с 15 декабря на 1 декабря. Отдельным решением станет перевод учреждений среднего профессионального образования на дистанционное обучение.
В настоящее время для этого готовится нормативная база.
Свердловским работодателям рекомендовали переводить непривитых на удаленку
https://ria.ru/20211102/covid-19-1757414995.html
Свердловским работодателям рекомендовали переводить непривитых на удаленку
Свердловским работодателям рекомендовали переводить непривитых на удаленку – РИА Новости, 02.11.2021
Свердловским работодателям рекомендовали переводить непривитых на удаленку
Свердловским работодателям рекомендовано перевести на удаленный режим работы всех, кто не вакцинировался хотя бы одним компонентом препарата от коронавируса,… РИА Новости, 02.11.2021
2021-11-02T17:12
2021-11-02T17:12
2021-11-02T17:12
общество
свердловская область
коронавирус в россии
/html/head/meta[@name=’og:title’]/@content
/html/head/meta[@name=’og:description’]/@content
https://cdnn21. img.ria.ru/images/07e5/07/1a/1742944710_0:0:3068:1727_1920x0_80_0_0_d2a65058c6360045fcac497997f10a60.jpg
img.ria.ru/images/07e5/07/1a/1742944710_0:0:3068:1727_1920x0_80_0_0_d2a65058c6360045fcac497997f10a60.jpg
ЕКАТЕРИНБУРГ, 2 ноя – РИА Новости. Свердловским работодателям рекомендовано перевести на удаленный режим работы всех, кто не вакцинировался хотя бы одним компонентом препарата от коронавируса, сообщил заместитель губернатора региона Павел Креков.Ранее вице-губернатор заявил, что в регионе может быть введен локдаун, если контрольные группы в течение следующих дней продолжат фиксировать нарушения режима QR-кодов в предприятиях торговли, общественного питания и других зонах, свободных от COVID-19.”Мы будем рекомендовать всем предприятиям и учреждениям, которые расположены на территории Свердловской области, перевести всех сотрудников, которые в настоящий момент не начали процесс вакцинации, на режим удаленной работы”, – сказал Креков.Также он уточнил, что власти сдвинут крайний срок получения второго компонента вакцины для допуска в зоны, свободные от COVID-19, с 15 на 1 декабря.По последним данным, всего в Свердловской области зарегистрированы 158,8 тысячи случаев заражения коронавирусом, в том числе 718 за последние сутки. Скончались 7,5 тысячи пациентов.
Скончались 7,5 тысячи пациентов.
https://ria.ru/20211102/lokdaun-1757388732.html
свердловская область
РИА Новости
7 495 645-6601
ФГУП МИА «Россия сегодня»
https://xn--c1acbl2abdlkab1og.xn--p1ai/awards/
2021
РИА Новости
7 495 645-6601
ФГУП МИА «Россия сегодня»
https://xn--c1acbl2abdlkab1og.xn--p1ai/awards/
Новости
ru-RU
https://ria.ru/docs/about/copyright.html
https://xn--c1acbl2abdlkab1og.xn--p1ai/
РИА Новости
7 495 645-6601
ФГУП МИА «Россия сегодня»
https://xn--c1acbl2abdlkab1og.xn--p1ai/awards/
https://cdnn21.img.ria.ru/images/07e5/07/1a/1742944710_0:0:2732:2048_1920x0_80_0_0_3adbba03b0506c6643daeb08b1bf4504.jpgРИА Новости
7 495 645-6601
ФГУП МИА «Россия сегодня»
https://xn--c1acbl2abdlkab1og. xn--p1ai/awards/
xn--p1ai/awards/
РИА Новости
7 495 645-6601
ФГУП МИА «Россия сегодня»
https://xn--c1acbl2abdlkab1og.xn--p1ai/awards/
общество, свердловская область, коронавирус в россии
Свердловским работодателям рекомендовали переводить непривитых на удаленку
ЕКАТЕРИНБУРГ, 2 ноя – РИА Новости. Свердловским работодателям рекомендовано перевести на удаленный режим работы всех, кто не вакцинировался хотя бы одним компонентом препарата от коронавируса, сообщил заместитель губернатора региона Павел Креков.
Ранее вице-губернатор заявил, что в регионе может быть введен локдаун, если контрольные группы в течение следующих дней продолжат фиксировать нарушения режима QR-кодов в предприятиях торговли, общественного питания и других зонах, свободных от COVID-19.
“Мы будем рекомендовать всем предприятиям и учреждениям, которые расположены на территории Свердловской области, перевести всех сотрудников, которые в настоящий момент не начали процесс вакцинации, на режим удаленной работы”, – сказал Креков.
Также он уточнил, что власти сдвинут крайний срок получения второго компонента вакцины для допуска в зоны, свободные от COVID-19, с 15 на 1 декабря.
По последним данным, всего в Свердловской области зарегистрированы 158,8 тысячи случаев заражения коронавирусом, в том числе 718 за последние сутки. Скончались 7,5 тысячи пациентов.
2 ноября, 15:12Распространение коронавирусаВ Свердловской области допустили введение локдаунаДистант продлят в вузах Москвы после 8 ноября / Интерфакс
В Российской академии народного хозяйства и госслужбы (РАНХиГС) сообщили “Интерфаксу”, что “в связи со сложной эпидемиологической ситуацией в столице и в соответствии с рекомендациями региональных властей руководством Академии было принято решение о продлении дистанционного формата обучения для студентов”.
“Учебный процесс будет организован в дистанционном формате до дальнейшего распоряжения”, – уточнили в Академии. Там добавили, что решение о формате обучения в филиалах вуза будет приниматься каждым регионом отдельно в зависимости от ситуации с COVID-19.
В Московском авиационном институте (МАИ) сообщили “Интерфаксу”, что с 8 по 13 ноября реализация основных образовательных программ будет организована исключительно с применением дистанционных образовательных технологий.
Аналогичным образом занятия будут проходить в Московском государственном университете им. Баумана. Согласно приказу ректора вуза, в этот период “первый прием задолженностей проводится в дистанционном формате, а приём задолженностей на аттестационных комиссиях – в очном, текущие контрольные будут сдаваться с применением онлайн-технологий”. Кроме того, ранее сообщалось, что “Бауманка” с 8 ноября вводит QR-коды для допуска на мероприятия вуза.
Такая же система “входного” контроля будет введена с 15 ноября для допуска на очные занятия студентов в Московском городском педагогическом университете (МГПУ). Для тех, кто не прошел вакцинацию от коронавируса и не имеет медицинского отвода, занятия будут проводиться только в дистанционном формате.
О переводе студентов на дистант после нерабочих дней сообщил ранее Национальный исследовательский университет “Высшая школа экономики”.
“С 8 ноября до конца 2021 года занятия в Вышке, в том числе сессия, будут проходить онлайн. Исключение сделано только для занятий, которые невозможно перенести полностью в дистанционный формат. Такие занятия будут проходить в малых группах до 15 человек при соблюдении необходимых мер”, – говорится в сообщении ВШЭ.
Между тем, творческие вузы, подведомственные Минкультуры РФ, надеются, что после окончания нерабочих дней с 8 ноября они вернутся к очному обучению. Пока распоряжений от ведомства о дальнейшем формате работы им не поступало.
“Пока такого решения нет. Вузы, конечно, могут работать дистанционно, мы будем гибкими, если ситуация с коронавирусом будет такова, что теоретические дисциплины можно будет вести онлайн, значит тогда так. А мастерство актера, сценическая речь и сценическое движение – нет, поэтому я надеюсь, что мы 8 ноября выйдем. Пока распоряжений министерства культуры нет”, – сообщил “Интерфаксу” ректор Высшего театрального училища (института) имени Щепкина Борис Любимов.
Ректор Театрального института им. Щукина Евгений Князев также сообщил, что пока решений не принято, но, если Минкультуры РФ подготовит соответствующий приказ, вуз будет проводить занятия со студентами дистанционно.
“Мы, если будет строгий приказ, будем работать дистанционно. Но театрам пока разрешено делать маленькие репетиции с теми людьми, у кого есть QR-коды и прививки. Мы, если нам дадут такое разрешение, мы и сейчас даем такое разрешение студентам, у которых есть QR-коды, они могут зайти в здание и заниматься самостоятельной работой, и дальше будем так работать. Те, кто не привился и у кого нет QR-кода, они в здание не допускаются и должны искать возможность для самостоятельной работы дома или в общежитии”, – сказал Князев агентству.
Всероссийский государственный институт кинематографии им.Герасимова также намерен после окончания нерабочих дней вернуться к очному обучению.
В РФ с 30 октября по 7 ноября указом президента объявлен период нерабочих дней, чтобы сбить волну распространения коронавируса, при этом руководителям регионов было дано право принимать решении об объявлении нерабочих дней до 30 октября или продлевать их после 7 ноября. В связи с указом президента Минобрнауки рекомендовало вузам перевести весь процесс обучения на этот период в онлайн или же предоставить студентам время для самостоятельной работы.
2 ноября вышел новый приказ ведомства, согласно которому высшие учебные заведения могут продолжить обучать студентов в дистанте и после 8 ноября. Промежуточная аттестация также может проводиться в дистанционном формате.
Читайте “Интерфакс-Образование” в “Facebook”, “ВКонтакте”, “Яндекс.Дзен” и “Twitter”
Страница не найдена
Новости
7 ноя
Губернатор Иркутской области Игорь Кобзев объявил о продлении каникул для школьников региона ещё на неделю.
3 ноя
Роспотребнадзор продлил действие санитарных правил по работе образовательных организаций в условиях распространения коронавирусной инфекции COVID-19.
1 ноя
У жительницы подмосковного Сергиева Посада сотрудники опеки увезли 14-летнюю дочь из школы в детский дом. Поводом для изъятия ребёнка из семьи послужила безобидная фотография. Девочка сделала селфи в ванной, полностью укутавшись в полотенце, а затем отправила фото однокласснику. Тот выложил его в соцсеть. По словам матери, классная руководительница посчитала фотографию слишком откровенной и сообщила о ней социальным службам. Через неделю, после вмешательства общественников и запроса RT в органы опеки, ребёнка вернули. Мать считает, что лица, ответственные за изъятие дочери, должны понести наказание.
31 окт
По меньшей мере четыре человека пострадали в результате вооружённого нападения на школу в столице Гаити Порт-о-Пренсе. Об этом сообщает агентство EFE.
30 окт
Выпускные экзамены в России в 2022 году пройдут в режиме, который действовал до 2020 года, если позволит эпидемиологическая ситуация, рассказали «Парламентской газете» в Рособрнадзоре и профильных комитетах обеих палат парламента.
30 окт
Выпускные экзамены в России в 2022 году пройдут в режиме, который действовал до 2020 года, если позволит эпидемиологическая ситуация, рассказали «Парламентской газете» в Рособрнадзоре и профильных комитетах обеих палат парламента.
29 окт
В Самаре бывшая учительница физкультуры, 24-летняя Анна Шмелёва (имя изменено. – RT), обвиняется в сексуальной связи с несовершеннолетней ученицей. В июне этого года мать школьницы (девочке сейчас 15 лет) написала заявление в Следственный комитет о том, что педагог якобы домогалась её дочери. СК возбудил уголовное дело по ч. 2 ст. 134 УК РФ («Мужеложство или лесбиянство с лицом, не достигшим шестнадцатилетнего возраста, совершённые лицом, достигшим восемнадцатилетнего возраста»). Адвокат Шмелёвой и её близкие считают, что родственники ученицы могли подать заявление в СК из мести: Шмелёва обращалась в опеку, чтобы семью взяли на контроль. В деталях громкого дела разбирался RT.
Контроль трансляции – обзор
1 Введение
Важным аспектом контроля экспрессии генов является диапазон скоростей деградации и трансляции различных мРНК. В последнее время начали накапливаться доказательства того, что контроль трансляции и деградации мРНК может вовлекать пару консервативных цитоплазматических гранул РНК (рис. 25.1). Один класс таких гранул РНК – это тело процессинга цитоплазмы, или P-тело. P-тельца представляют собой динамические агрегаты нетранслирующих мРНК в сочетании с репрессорами трансляции и белками, участвующими в деаденилировании, декапировании и экзонуклеолитическом распаде от 5 ‘до 3’ (Parker and Sheth, 2007).P-тела и собранные в них мРНП представляют интерес по нескольким причинам. Они участвуют в репрессии трансляции (Coller and Parker, 2005; Holmes et al. , 2004), нормальном распаде мРНК (Cougot et al., 2004; Sheth and Parker, 2003), распаде, опосредованном нонсенсом (Sheth and Parker, 2006; Unterholzner and Izaurralde, 2004), miRNA-опосредованная репрессия у многоклеточных животных (Liu et al. , 2005; Pillai et al. , 2005) и хранение мРНК (Bhattacharyya et al., 2006; Brengues et al. , 2005). Как минимум, Р-тельца служат маркерами, которые пропорциональны концентрации мРНП в комплексе с механизмом репрессии распада / трансляции мРНК и могут иметь дополнительные биохимические свойства, которые влияют на контроль трансляции и / или деградации мРНК.
Рисунок 25.1. Р-тела и стресс-гранулы при различных условиях роста и индукции. (A) Mid-log клетки дикого типа (yRP840), трансформированные pRP1660. Dcp2-mCh служит маркером P-body. Примечание : Очень слабые и большие фокусы в столбце P-тела (A и B) представляют собой фактическую вакуолярную аутофлуоресценцию. (B) Mid-log клетки дикого типа (yRP840), трансформированные pRP1660 и подвергшиеся 10 мин-стрессу депривации Glu. Dcp2-mCh служит маркером P-body. (C) клетки дикого типа с высокой OD (BY4741), трансформированные pRP1657; 2 дня роста в минимальной среде. Edc3-mCh служит маркером P-body. Примечание : Фокусы Pab1 с высоким OD не могут быть прямо эквивалентны гранулам среднего логарифмического напряжения (см. Основной текст).
Стресс-гранулы представляют собой вторую гранулу мРНП, участвующую в контроле трансляции, и были широко изучены в клетках млекопитающих; обзоры см. в Anderson and Kedersha (2006, 2008). Стресс-гранулы обычно не наблюдаются в нормальных условиях роста в клетках дрожжей или млекопитающих и значительно увеличиваются в ответ на дефекты инициации трансляции, включая снижение функции eIF2 или eIF4A (Dang et al. , 2006; Kedersha et al. , 2002 ; Mazroui et al., 2006). Поскольку стрессовые реакции часто включают временное ингибирование инициации трансляции, стрессовые гранулы накапливаются во время широкого диапазона стрессовых реакций. Утверждается, что стрессовые гранулы функционируют как центры «сортировки» мРНК, покидающих полисомы во время стресса, где мРНК либо сортируются в Р-тельца для распада, поддерживаются в хранящемся нетранслируемом состоянии или возвращаются для трансляции (Anderson and Kedersha, 2006, 2008). ).
Недавние результаты показали, что гранулы мРНП, подобные стрессовым гранулам млекопитающих, могут образовываться у почкующихся дрожжей.Это было впервые предположено на основании наблюдения, что факторы инициации трансляции eIF4E, eIF4G и Pab1p, компоненты стрессовых гранул млекопитающих, образуют в дрожжах очаги во время депривации глюкозы и в условиях высокой OD, которые могут либо совместно локализоваться с Р-тельцами, либо отличаться от них ( Бренге и Паркер, 2007; Хойл и др. , 2007). Эти стрессовые гранулы, также называемые тельцами EGP, также содержат мРНК (Hoyle et al. , 2007). Дополнительным доказательством того, что эти тельца EGP или дрожжевые стрессовые гранулы эквивалентны стрессовым гранулам млекопитающих, является то, что они содержат дрожжевые ортологи нескольких белков, обнаруженных в стрессовых гранулах млекопитающих (Таблица 25.2) и используют аналогичные правила сборки (Buchan et al. , 2008). Такие правила сборки включают требование нетранслирующей мРНК, стимуляцию сниженными функциональными уровнями фактора инициации трансляции eIF2 и требование сходных факторов сборки белка (Buchan et al. , 2008).
Р-тельца и стрессовые гранулы взаимодействуют и часто состыковываются в клетках млекопитающих, тогда как у почкующихся дрожжей они преимущественно перекрываются (Buchan et al. , 2008; Kedersha et al., 2005). Бутонированные дрожжи представляют собой хорошую систему для анализа взаимодействий Р-тельца и стрессовых гранул, поскольку сборку обеих гранул можно предотвратить или модифицировать в различных мутантных штаммах (Buchan et al., 2008; Coller and Parker, 2005; Decker et al. , 2007; Тейшейра, Паркер, 2007). Действительно, такие эксперименты показали, что образование стрессовых гранул в некоторых случаях усиливается уже существующими Р-тельцами, предполагая функциональную взаимосвязь между ними.
Анализ оборота мРНК и репрессии трансляции может включать изучение аспектов состава и сборки как P-телец, так и стрессовых гранул, учитывая концентрацию мРНК, факторы распада, репрессоры трансляции и факторы инициации в этих гранулах.В этой главе мы описываем методы анализа Р-тел и стрессовых гранул у почкующихся дрожжей, Saccharomyces cerevisiae . Мы сосредоточены на описании методов, чтобы ответить на три общих вопроса: (а) Накапливается ли данный белок или мРНК в Р-тельцах или стрессовых гранулах? (b) Изменяет ли конкретное возмущение (например, мутация, сверхэкспрессия или сигнал окружающей среды) качественно размер или количество Р-телец или стрессовых гранул? (c) Имеется ли поддающееся количественной оценке изменение количества и размера Р-тел или стрессовых гранул в данной популяции клеток?
Контроль трансляции и miRNA-зависимой репрессии с помощью нового поли (A) связывающего белка, hnRNP-Q
Образец цитирования: Свиткин Ю.В., Янагия А., Каретников А.Е., Ален Т., Фабиан М.Р., Хоуторский А. и др.(2013) Контроль трансляции и miRNA-зависимой репрессии с помощью нового поли (A) связывающего белка, hnRNP-Q. PLoS Biol 11 (5): e1001564. https://doi.org/10.1371/journal.pbio.1001564
Академический редактор: Йенс Ликке-Андерсен, Калифорнийский университет, Сан-Диего, США
Поступила: 15 февраля 2013 г .; Одобрена: 10 апреля 2013 г .; Опубликован: 21 мая 2013 г.
Авторские права: © 2013 Svitkin et al.Это статья в открытом доступе, распространяемая в соответствии с условиями лицензии Creative Commons Attribution License, которая разрешает неограниченное использование, распространение и воспроизведение на любом носителе при условии указания автора и источника.
Финансирование: Это исследование было поддержано грантом GM66157 Национального института здравоохранения США и грантом MOP-7214 Канадского института медицинских исследований (CIHR) NS. AEK является стипендиатом постдокторской стипендии CIHR. IT имеет награду CIHR Young Investigator Award.Финансирующие организации не играли никакой роли в дизайне исследования, сборе и анализе данных, принятии решения о публикации или подготовке рукописи.
Конкурирующие интересы: Авторы заявили об отсутствии конкурирующих интересов.
Сокращения: ВГС, вирус гепатита С; hnRNP-Q, гетерогенный ядерный рибонуклеопротеин Q; IRES, внутренний сайт входа рибосомы; ПАБП, связывающий поли (A) белок; Paip2, Белок 2, взаимодействующий с PABP; PV, полиовирус; РРЛ, лизат ретикулоцитов кролика; SYNCRIP, синаптотагмин-связывающий, цитоплазматический белок, взаимодействующий с РНК
Введение
Белки, которые образуют динамические мультипротеиновые комплексы с эукариотическими мРНК, играют важную роль в контроле экспрессии генов [1].Состав и архитектура частиц рибонуклеопротеина-мессенджера (мРНП) в значительной степени определяет их распределение между различными субклеточными структурами (т.е. полисомами, стрессовыми гранулами или процессинговыми телами) и, в конечном итоге, скоростью трансляции и деградации мРНК [2]. Недавний анализ выявил неожиданно широкий набор мРНК-связывающих белков млекопитающих с в значительной степени неизвестными функциями [3]. Во время инициации трансляции, которая является лимитирующей и наиболее регулируемой стадией синтеза белка, рибосома 80S рекрутируется на мРНК и позиционируется в кодоне инициации [4].Этому процессу способствует связывание эукариотического фактора инициации 4E (eIF4E) с кэп-структурой m 7 G на 5′-конце мРНК и поли (A) связывающим белком (PABP) с 3′-поли (A) хвост. Стимулирующие эффекты кэп-структуры и поли (А) -хвоста на трансляцию синергетичны. eIF4E является субъединицей комплекса eIF4F, который также включает eIF4A, РНК-зависимую АТФазу / РНК-геликазу, и eIF4G, высокомолекулярный каркасный белок [5]. eIF4G взаимодействует с PABP [6] – [8] и eIF3, который является мостиком между eIF4G и 40S субъединицей рибосомы [9].Эти взаимодействия циркулируют мРНК [10] и усиливают трансляцию (обзор в [11] – [13]).
PABP – это высоко эволюционно консервативный белок, который был впервые описан, насколько нам известно, четыре десятилетия назад [14]. Он содержит четыре мотива распознавания РНК (RRM) и богатую пролином С-концевую область, которая участвует в межбелковых взаимодействиях. PABP связывает поли (A) кооперативным образом и с периодичностью ~ 27 нуклеотидов [15]. PABP, ассоциированный с 3′-концом, является критическим детерминантом трансляционной активности мРНК, которая действует в цис- [16].Истощение PABP из трансляционных экстрактов снижает связывание eIF4E со структурой кэпа и резко ингибирует образование комплекса инициации 48S и 80S рибосом [17]. PABP также может играть роль на поздней стадии инициации, способствуя присоединению рибосомных субъединиц [18], а также во время терминации и рециклинга рибосом путем образования комплекса с eRF3 [19]. Эти результаты подчеркивают важность PABP для глобального перевода. Выполняя другую роль, PABP усиливает ассоциацию индуцированного микроРНК комплекса молчания (miRISC) со специфическими мРНК для усиления репрессии трансляции, опосредованной miRNA [20].Наконец, PABP регулирует деаденилирование мРНК, которое является первым и обычно лимитирующим этапом деградации мРНК [21]. Мы и другие недавно показали, что miRISC, который включает белки Argonaute (AGO) и GW182 [22] – [24], связывает PABP (через GW182) и привлекает деаденилазу CNOT7 / CAF1, чтобы способствовать укорочению поли (A) хвоста [25], [26]. Эти и другие исследования указывают на участие PABP в мРНК-специфической регуляции синтеза белка [27]. Интересно, что PABP является предметом посттрансляционных модификаций, функциональное значение которых еще предстоит установить [28].
Два белка, взаимодействующих с PABP, Paip1 и Paip2, модулируют активность PABP при трансляции. Paip1 – положительный регулятор трансляции [29], [30]. Напротив, Paip2 ингибирует трансляцию, вытесняя PABP из поли (A) хвоста и eIF4G [31], [32]. Ожидается, что диссоциация PABP от поли (A) хвоста приведет к ремоделированию мРНП. Здесь мы сообщаем о взаимодействии изоформы 2 гетерогенного ядерного рибонуклеопротеина Q мыши (в дальнейшем называемой hnRNP-Q2) с поли (A) хвостом. Мы также показываем, что hnRNP-Q2 конкурирует с PABP за связывание поли (A), чтобы ингибировать глобальный синтез белка как in vitro, так и in vivo, и ослаблять опосредованную miRNA репрессию мРНК.Эти данные указывают на участие hnRNP-Q2 в контроле многофункциональной активности PABP.
Результаты
Paip2-индуцированное ремоделирование mRNP
Paip2 резко снижает сродство PABP к поли (А) в модельных системах, содержащих поли (А) и чистые рекомбинантные белки [31]. Чтобы изучить влияние Paip2 на образование комплекса РАВР-поли (A) в более физиологических условиях, то есть в контексте клеточных экстрактов, мы использовали УФ-индуцированное перекрестное связывание, которое является надежным методом обнаружения специфических взаимодействий РНК-белок [ 33].Для этого анализа мРНК кроличьего глобина удлиняли по 3′-концу с использованием [α- 32 P] АТФ и дрожжевой поли (A) полимеразы. МРНК инкубировали с обработанным микрококковой нуклеазой лизатом ретикулоцитов кролика (RRL), бесклеточным экстрактом S10 HeLa или Кребса, подвергали УФ-облучению и расщепляли смесью РНКаз. Белки, связанные с поли (A) хвостом, анализировали с помощью SDS-PAGE и авторадиографии. Во всех экстрактах наблюдали почти исключительное сшивание полипептида ∼70 кДа (p70) с поли (A) (рис. 1A, B, C; дорожка 1).Добавление Paip2 предотвращает перекрестное связывание p70, указывая на диссоциацию комплекса p70-poly (A) (рис. 1A, B, C; дорожка 2). Эти результаты убедительно свидетельствуют о том, что p70 является PABP. Действительно, p70 не был обнаружен в экстрактах, лишенных PABP с использованием аффинной матрицы GST-Paip2 (рис. 1A, B, C; дорожка 3) [34]. Неожиданно мы наблюдали связывание новых белков (p68 и p58 / 59) с поли (A) хвостом после секвестрации PABP с помощью Paip2 или истощения PABP (рис. 1A, B, C; дорожки 2, 3). Дополнение экстрактов, истощенных по PABP, рекомбинантным PABP устраняет связывание этих белков с поли (A) (рис. 1A, B, C; дорожка 4).Интересно, что p68 проявляется в виде единственной заметной полосы в RRL, которая была дополнена Paip2 или лишена PABP (рис. 1A, дорожки 2, 3). p68 также обнаруживался в контрольном RRL, хотя и в виде слабой полосы (рис. 1A, дорожка 1). Следовательно, мы хотели идентифицировать этот белок.
Рис. 1. Обнаружение белков, взаимодействующих с поли (А), с помощью УФ-сшивки. Цитоплазматические экстракты
RRL (A), HeLa (B) или Krebs (C) S10 подвергали УФ-индуцированному перекрестному связыванию с хвостом 32 P-poly (A).Экстракты, которые были либо ложно истощены (контроль S10), либо лишены PABP, инкубировали с 32 P поли (A) мРНК глобина, меченной хвостом, при 32 ° C в течение 10 мин. Перед добавлением мРНК реакционные смеси предварительно инкубировали при 32 ° C в течение 2 минут с GST-Paip2 (20 мкг / мл) или PABP (5 мкг / мл), как указано. После УФ-облучения и обработки РНКазой меченые белки анализировали с помощью SDS-PAGE и авторадиографии. Справа указаны позиции маркеров молекулярной массы. Вестерн-блоттинг с антителом против PABP использовали для подтверждения достаточного истощения PABP из экстрактов (репрезентативные анализы см. [54]).Следует отметить, что в этом и других анализах сшивания УФ-светом PABP выглядел как суетливая полоса, скорее всего, потому, что расщепление РНКазой не дает гомогенных поперечно-сшитых фрагментов РНК.
https://doi.org/10.1371/journal.pbio.1001564.g001
Идентификация p68 как hnRNP-Q
Для определения идентичности p68 в клетках HeLa цитоплазматический экстракт HeLa был лишен PABP и подвергнут хроматографии на поли (A) -сефарозе. После промывки шариков буфером, содержащим 0.2 M KCl, взаимодействующие с поли (A) белки последовательно элюировали буферами, содержащими 1 M KCl и 2 M LiCl, и разделяли с помощью SDS-PAGE (рисунок S1A). В отличие от УФ-сшивания, этот анализ захватил многие белки, скорее всего, потому, что многие из них связываются с сахарно-фосфатным остовом РНК или других РНК-связывающих белков. В соответствии с ролью ионных РНК-белковых взаимодействий, характер связывания поли (A) белков в более жесткой промывке (2 M LiCl) был менее сложным, чем при промывке 1 M KCl.Два кандидата на р68 (полипептиды ~ 69 кДа и ~ 70 кДа; полосы 1 и 2 соответственно) вырезали из геля, расщепляли трипсином и анализировали масс-спектрометрией. Один белок в полосе 1 был идентифицирован как hnRNP-Q (номер доступа UniProtKB O60506) с 36,3% -ным покрытием последовательности 22 уникальными пептидами (таблица S1). Полоса 2 была идентифицирована как родственный белок теплового шока 71 кДа (HSP7C, UniProtKB, инвентарный номер P11142) с 50,3% покрытием последовательности 24 уникальными пептидами (неопубликованные данные). Чтобы определить, является ли какой-либо из двух белков поли (A) -сшиваемым p68, мы выполнили иммунопреципитацию p68 из RRL после УФ-индуцированного перекрестного сшивания.Моноклональное антитело против hnRNP-Q (18E4) эффективно осаждает 32 P-меченного p68 (фигура S1B). Напротив, два антитела против hsp70, а также антитело против PABP не смогли этого сделать. Белок p68 экстракта Кребса также осаждали антителом против hnRNP-Q (неопубликованные данные). Эти результаты демонстрируют, что p68 идентичен hnRNP-Q.
hnRNP-Q, также называемый NS1-ассоциированным белком-1 (NSAP1) [35], является обильным и повсеместно экспрессируемым белком [36], которому приписаны функции в сплайсинге пре-мРНК и метаболизме мРНК [37] – [40] ], а также роль в IRES-опосредованной трансляции [41] – [45].hnRNP-Q очень гомологичен hnRNP-R и содержит N-концевой кислотный домен, три RRMs и богатую RGG C-концевую область, которая может участвовать в связывании РНК и межбелковых взаимодействиях [37]. Множественные изоформы hnRNP-Q (семь у человека и две у мыши) являются производными от альтернативного сплайсинга одного гена [37]. Посттрансляционные модификации hnRNP-Q, которые включают фосфорилирование и метилирование, могут определять его субклеточную локализацию и свойства связывания РНК [46], [47]. У мышей небольшой (длиной 562 аминокислоты) вариант сплайсинга hnRNP-Q, называемый SYNaptotagmin-связывающим цитоплазматическим РНК-взаимодействующим белком (SYNCRIP) или изоформой 2 hnRNP-Q (hnRNP-Q2; номер доступа NP_062770.1), в основном цитоплазматический, тогда как более длинная изоформа 1 hnRNP-Q находится в ядре [40], [46], [48]. Ортологом человеческого hnRNP-Q2 мыши является изоформа 6 hnRNP-Q (hnRNP-Q6; регистрационный номер NP_001153149.1), последовательность которой идентична последовательности hnRNP-Q2, за исключением аланина вместо серина в положении 357. В частности, цитоплазматический изоформы hnRNP-Q содержат один, а не два сигнала ядерной локализации [37]. Мы обнаружили, что RRL содержит единственную изоформу hnRNP-Q, которая мигрирует вместе с самой маленькой изоформой hnRNP-Q клеток Кребса или клеток HeLa (рисунок S1C).Чтобы определить, связана ли цитоплазматическая изоформа hnRNP-Q с активно транслируемыми полисомными мРНК или неактивными мРНП, цитоплазматический экстракт HeLa центрифугировали в градиенте плотности сахарозы. Белки из каждой фракции анализировали вестерн-блоттингом с использованием антител против hnRNP-Q и PABP. Примечательно, что, несмотря на общую высокую аффинность связывания с поли (А), hnRNP-Q и PABP различались в отношении их субклеточного распределения. HnRNP-Q присутствовала во фракциях несвязанный белок / свободный мРНП (вместе с eIF4A, eIF4E и Paip2), а также во фракции 40S рибосомной субъединицы, но не во фракциях полисом (Рисунок S2).Напротив, PABP и белок упаковки мРНК YB-1 [49] связаны с полисомами в дополнение к их присутствию во фракциях верхнего градиента. Эти результаты указывают на ассоциацию hnRNP-Q с нетранслируемыми мРНП и согласуются с другими исследованиями субклеточного распределения NSAP1 / hnRNP-Q, PABP и YB-1 [42], [45], [50], [51] ].
Конкуренция между hnRNP-Q2 и PABP за поли (A) связывание
Интересно, что сообщалось, что SYNCRIP / hnRNP-Q2 проявляет преимущественное связывание с поли (A) [48].Следовательно, мы хотели изучить специфичность связывания поли (A) hnRNP-Q2, выполнив эксперименты по конкуренции РНК в RRL. Добавление поли (A) (5 мкг / мл) к RRL, обедненному PABP, полностью ингибировало УФ-сшивание hnRNP-Q2 с поли (A) хвостом (рис. 2A). Поли (G), поли (U) и 18S рибосомная РНК (рРНК) не влияла на сшивание hnRNP-Q2, в то время как поли (C) оказывала слабое ингибирующее действие. Поли (A) также специфически ингибировал поперечное сшивание PABP с поли (A) хвостом, как это было определено в контрольном (ложном) RRL.Таким образом, внутри RRL hnRNP-Q2 проявляет предпочтение по отношению к поли (A).
Рисунок 2. Специфичность связывания hnRNP-Q2 с поли (A) хвостом.
(A) Белки контрольного или обедненного PABP RRL УФ, поперечно сшитые с P-поли (A) хвостом 32 , анализировали с помощью SDS-PAGE, как описано для Фиг.1. Конкуренты РНК, 18S рРНК, поли (A), поли (G), поли (U) и поли (С) включали в реакционные смеси в концентрациях 5 мкг / мл. (B) Рекомбинантный hnRNP-Q2 (2 мкг) анализировали с помощью SDS-PAGE и окрашивания кумасси синим.Предварительно окрашенные маркеры молекулярной массы (MBI Fermentas) показаны в правой полосе. (C) Высокоаффинное связывание hnRNP-Q2 с олиго (A). EMSA выполняли, как описано в разделе «Материалы и методы». Для каждой дорожки инкубировали постоянное небольшое количество РНК 32 P-oligo (A) 30 с указанными концентрациями hnRNP-Q2. Значение K d ∼30 нМ было рассчитано на основе трех независимых экспериментов.
https://doi.org/10.1371/journal.pbio.1001564.g002
Чтобы количественно охарактеризовать взаимодействие hnRNP-Q2 / поли (A), бактериально экспрессируемый His-меченный мышиной hnRNP-Q2 подвергали аффинной очистке с помощью Ni 2+ -NTA агарозной хроматографии. Анализ SDS-PAGE и УФ-спектра показал, что препарат hnRNP-Q2 практически не содержит загрязняющих белков и нуклеиновых кислот (рис. 2B и неопубликованные данные). Рекомбинантный hnRNP-Q2 использовали в анализе сдвига электрофоретической подвижности (EMSA), в котором постоянное небольшое количество 5 ‘ 32 P-меченного олиго (A 30 ) титровали возрастающими количествами hnRNP-Q2.После инкубации комплексы РНК / белок были отделены от свободного олигонуклеотида (A 30 ) с помощью нативного гель-электрофореза и количественно определены с помощью фосфорного сканера (рис. 2С). Кажущееся K d для hnRNP-Q2 (что равно концентрации белка, при которой 50% зонда превращается в комплекс), по оценкам, составляло ~ 30 нМ. Интересно, что количество сдвинутого зонда постепенно увеличивалось с увеличением количества hnRNP-Q2, указывая на то, что образование стабильного комплекса требует кооперативного связывания hnRNP-Q2.Таким образом, hnRNP-Q2 активно связывается с олиго (A), хотя и менее сильно, чем PABP, для которого сообщалось о K d, равном 4-7 нМ [52], [53].
Чтобы определить, конкурирует ли hnRNP-Q2 с PABP за связывание поли (A), его удаляли из RRL с помощью антитела против hnRNP-Q. Вестерн-блоттинг выявил эффективное (~ 90%) истощение hnRNP-Q2, но не β-актина, который служил контролем нагрузки (рис. 3A). В отсутствие эндогенного hnRNP-Q2 происходило эффективное перекрестное связывание PABP с поли (A) хвостом глобиновой мРНК (рис. 3B).Добавление увеличивающихся количеств hnRNP-Q2 постепенно уменьшало сшивание PABP. В обратном эксперименте к RRL с истощенным PABP [54] добавляли возрастающие концентрации рекомбинантного PABP. В отсутствие PABP hnRNP-Q2 по умолчанию связывался поли (A) хвостом, проявляясь в виде заметной полосы (рис. 3C). Полоса hnRNP-Q2 постепенно исчезла, в то время как полоса PABP усилилась при добавлении к реакционной смеси возрастающих концентраций PABP. Эти результаты ясно демонстрируют, что PABP и hnRNP-Q2 конкурируют друг с другом за связывание поли (A).
Рисунок 3. Конкуренция между hnRNP-Q2 и PABP за связывание с поли (A) хвостом в RRL.
(A) Вестерн-блот-анализ RRL, истощенного либо анти-FLAG (контрольный RRL), либо анти-hnRNP-Q антителом. Блот анализировали на наличие анти-hnRNP-Q или анти-β-актина (контроль нагрузки). (B) Белки контрольного или обедненного hnRNP-Q2 RRL, которые перекрестно связываются с P-поли (A) хвостом 32 в присутствии указанных концентраций рекомбинантного hnRNP-Q2. (C) Белки контроля или RRL с обедненным PABP, которые перекрестно связываются с P-поли (A) хвостом 32 в присутствии указанных концентраций рекомбинантного PABP.
https://doi.org/10.1371/journal.pbio.1001564.g003
HnRNP-Q2 является ингибитором PABP / Poly (A) -зависимой трансляции
Показав, что hnRNP-Q2 и PABP конкурируют за связывание с поли (A) хвостом, мы предсказали, что hnRNP-Q2 будет противодействовать активности PABP при трансляции. Чтобы исследовать это, эндогенный hnRNP-Q2 был иммунодеплетирован из экстракта Кребса (истощение ~ 90%; фиг. 4A). Истощение hnRNP-Q2 стимулировало трансляцию кэпированной и полиаденилированной (A 98 ) мРНК люциферазы [обозначенной как Cap-Luc-p (A) 98 мРНК] на ~ 3 раза.5-кратный (Рисунок 4B). Стимулирующий эффект истощения hnRNP-Q2 на трансляцию не был обусловлен совместным истощением YB-1 (рис. 4A), белка, упаковывающего мРНК, и общего репрессора трансляции [49], [54], [55]. Добавление hnRNP-Q2 обратно к истощенному экстракту уменьшало трансляцию, и это ингибирование было зависимым от дозы hnRNP-Q2. Примечательно, что количества hnRNP-Q2, добавленные в этом и других анализах, находились в диапазоне концентраций, обычно обнаруживаемых в экстрактах Кребса (~ 30 мкг / мл; Рисунок S3). Чтобы исключить возможность того, что hnRNP-Q2 ингибирует синтез белка путем дестабилизации мРНК, в экстрактах трансляции, содержащих или не содержащих hnRNP-Q2, отслеживали распад 32 P-меченой мРНК Cap-Luc-p (A) 98 .Cap-Luc-p (A) 98 мРНК оставалась интактной в контрольных и истощенных hnRNP-Q2 трансляционных экстрактах в течение 2-часового периода инкубации (рис. 4C). Кроме того, добавление hnRNP-Q2 (30 мкг / мл) к обедненному экстракту не оказало влияния на стабильность мРНК Cap-Luc-p (A) 98 .
Рисунок 4. HnRNP-Q2-опосредованное ингибирование трансляции в экстрактах Кребса.
(A) Вестерн-блоттинг экстрактов Кребса S10, обедненных антителами против FLAG (контроль S10) или против hnRNP-Q. Блот анализировали на hnRNP-Q, YB-1 или β-актин (контроль нагрузки).(B) Мок-истощенные (контрольные) или истощенные по hnRNP-Q2 экстракты S10 Кребса были запрограммированы с помощью мРНК Cap-Luc-p (A) 98 в отсутствие или в присутствии указанных концентраций hnRNP-Q2. (C) Стабильность Cap-Luc-p (A) 98 мРНК. 32 P-меченный Cap-Luc-p (A) 98 использовали для программирования ложно-истощенных (контрольных) или истощенных по hnRNP-Q2 экстрактов S10 Krebs, не дополненных или дополненных рекомбинантным hnRNP-Q2 (30 мкг / мл. ), как указано. Полную РНК выделяли в указанные моменты времени из аликвот реакционной смеси, разделяли электрофорезом в формальдегид-агарозном геле и переносили на мембрану.Cap-Luc-p (A) 98 мРНК и 28S рРНК были обнаружены авторадиографией (левая панель, вверху) и окрашиванием (левая панель, внизу), соответственно. Cap-Luc-p (A) 98 интенсивности полос мРНК определяли и скорректировали с учетом нагрузки 28S рРНК (правая панель). Значения для мРНК Cap-Luc-p (A) 98 , выделенной в начале инкубации, были приняты за 100%. (D и E) Контрольный (ложно истощенный) и экстракты Кребса, обедненные hnRNP-Q2, были запрограммированы с PV IRES-Luc-p (A) 98 (D) или HCV IRES-Luc-p (A) 98 ( E) мРНК.hnRNP-Q2 добавляли к реакционным смесям в указанных концентрациях. На панелях B, D и E данные представляют собой средние значения трех анализов со стандартными отклонениями от среднего.
https://doi.org/10.1371/journal.pbio.1001564.g004
Затем мы исследовали влияние hnRNP-Q2 на кэп-независимую трансляцию, управляемую различными вирусными внутренними сайтами связывания рибосом (IRES). Экстракт, обедненный hnRNP-Q2, был в ~ 2,2 раза более активен, чем экстракт ложного истощения, в поддержке трансляции из IRES полиовируса (PV) (рис. 4D).Однако трансляция IRES вируса гепатита C (HCV) (которая не зависит от PABP и eIF4G, в отличие от PV IRES) не была значительно усилена истощением hnRNP-Q (рис. 4E). В соответствии с этими результатами в экстракте, обедненном hnRNP-Q2, PV IRES проявлял большую чувствительность к ингибированию рекомбинантным hnRNP-Q2 по сравнению с IRES HCV (фиг. 4, сравните панели D и E). Таким образом, конкуренция со стороны hnRNP-Q2 существенно не влияет на функцию рибосом и факторов трансляции, кроме комплекса PABP / eIF4G.
HnRNP-Q ингибирует рекрутирование eIF4F на мРНК
Чтобы выяснить, нацелена ли конкуренция со стороны hnRNP-Q2 на этап инициации трансляции, мы исследовали связывание рибосом с использованием коммерчески обработанных нуклеазами RRL. Хотя зависимость RRL от кэп- и поли (A) хвоста уменьшается после обработки нуклеазой [56], [57], значительная зависимость от этих структур для трансляции наблюдается при низких уровнях входящей мРНК и повышенных концентрациях ионов калия (Рисунок S4). [54], [58] – [60].Например, при дополнительной концентрации KCl 60 мМ кэппирование и полиаденилирование усиливают трансляцию мРНК Luc (0,5 мкг / мл) в 12,5 и 3,3 раза соответственно (рис. S4B, D). Поэтому нижеприведенные анализы проводили с использованием RRL с добавлением KCl (60 мМ) и низких (<0,5 мкг / мл) концентраций мРНК. Чтобы исследовать образование комплекса инициации 80S рибосомы, RRL инкубировали с радиоактивно меченной мРНК глобина в присутствии циклогексимида. Комплекс 80S отделяли от несвязанной мРНК центрифугированием в градиенте сахарозы.Добавление hnRNP-Q2 (20 мкг / мл) к контрольным или обедненным hnRNP-Q2 RRL ингибировало образование комплекса инициации 80S в 2,3–3 раза (фигуры 5A, B). Аналогичное снижение рекрутирования 80S рибосом в присутствии hnRNP-Q2 наблюдалось в нормальных или обедненных hnRNP-Q2 экстрактах Кребса (рис. S5). Чтобы определить, нацелен ли hnRNP-Q2 также на образование 48S пре-инициаторного комплекса, соединение 60S рибосомных субъединиц ингибировалось GMPPNP, негидролизуемым аналогом GTP [17]. В RRL, дополненном GMPPNP, меченая мРНК перераспределялась из фракций 80S градиента во фракции 48S, тем самым подтверждая анализ (рис. 5C).Важно отметить, что добавление hnRNP-Q2 (24 мкг / мл) к RRL, истощенному по hnRNP-Q2, ингибировало образование 48S инициирующего комплекса примерно в 5 раз с глубоким сдвигом мРНК во фракции RNP (рис. 5D). Чтобы определить, ингибирует ли hnRNP-Q2 трансляцию до образования комплекса 48S, мы исследовали взаимодействие eIF4E с кэп-структурой в ложных и обедненных hnRNP-Q2 RRL путем химического сшивания лизатов с полиаденилированной мРНК Luc 32 P-меченой на 5-дюймовой крышке. Ранее мы показали, что этот анализ обеспечивает высоконадежную оценку активности eIF4F [54].В истощенных hnRNP-Q2 RRL перекрестное связывание eIF4E было усилено в ~ 1,5 раза по сравнению с ложно-истощенными RRL (фиг. 5E). Добавление увеличивающихся концентраций hnRNP-Q2 уменьшало сшивание дозозависимым образом (до 25% от контроля). Таким образом, hnRNP-Q2 нарушает взаимодействие eIF4E с кэп-структурой. Поскольку PABP стимулирует взаимодействие eIF4E-cap [17], [61], наиболее вероятно, что hnRNP-Q2 действует, ингибируя эту функцию PABP. Чтобы получить доказательства того, что hnRNP-Q2 нацелен на факторы группы eIF4 в экстракте Кребса, в эту систему были добавлены очищенные eIF4F, eIF4A, eIF4E и eIF4B, которые либо лишены, либо содержат hnRNP-Q2.Эти факторы стимулировали трансляцию мРНК Cap-Luc-p (A) 98 , что согласуется с их присутствием в ограниченных количествах в экстрактах Кребса (рис. S6 и [62]). В соответствии с частичной репрессией активности eIF4F с помощью hnRNP-Q2, экзогенный eIF4F снимал опосредованное hnRNP-Q2 ингибирование трансляции (от 5,5 до 1,4 раза). eIF4A, eIF4E и eIF4B также снимали ингибирование трансляции hnRNP-Q2, хотя и менее эффективно, чем eIF4F.
Рис. 5. HnRNP-Q2 ингибирует распознавание структуры G-кэпа с помощью факторов инициации трансляции.
(A – D) Ингибирование образования инициирующего комплекса 80S и 48S с помощью hnRNP-Q2 в обработанных нуклеазой RRL. Связывание 80S рибосомы с 3′-концом меченой мРНК глобина анализировали в RRL, дополненном циклогексимидом (0,6 мМ), нормальном (A) или обедненном hnRNP-Q2 (B), в присутствии контрольного буфера (квадраты) или рекомбинантного hnRNP- Q2 (15 мкг / мл) (треугольники). (C) Подтверждение образования комплекса 48S до инициации в присутствии GMPPNP. GTP или GMPPNP добавляли к реакционным смесям при конечной концентрации 2 мМ, как указано.Другие условия были аналогичны тем, которые описаны для панели B. (D) Формирование прединициативного комплекса 48S в истощенных hnRNP-Q2 RRL в присутствии GMPPNP и либо контрольного буфера (квадраты), либо hnRNP-Q2 (25 мкг / мл) ( треугольники). Реакционные смеси анализировали на градиентах сахарозы 5 мл 15–30% (A и B) или 11 мл 10–30% (C и D). (E) HnRNP-Q2 дозозависимое ингибирование связывания eIF4E с кэп-структурой m 7 G в RRL, как было проанализировано с помощью химического сшивания. Контрольный и истощенный hnRNP-Q2 RRL инкубировали с окисленной 32 P-cap-меченной поли (A) -расширенной мРНК Luc в отсутствие или в присутствии указанных концентраций рекомбинантного hnRNP-Q2.Указаны положения eIF4E и eIF4A. Относительные эффективности сшивания eIF4E указаны внизу (значение, полученное для контрольного RRL, было установлено за 100%).
https://doi.org/10.1371/journal.pbio.1001564.g005
Длинные поли (A) хвосты усиливают репрессию трансляции, опосредованную hnRNP-Q2
Длина поли (A) хвоста определяет количество молекул PABP, связанных с мРНК, тем самым косвенно контролируя PABP-зависимую трансляцию.Чтобы изучить, зависит ли опосредованное hnRNP-Q2 ингибирование трансляции от длины поли (A) хвоста, мы сравнили влияние hnRNP-Q2 на трансляцию мРНК Luc либо без (A 0 ), либо с поли (A). (A) хвост увеличивающейся длины (A 15 , A 30 , A 45 , A 90 и A 250 ). Для этих исследований мы использовали экстракт Кребса, который не подвергался обработке нуклеазами. В нескольких исследованиях было доказано, что отсутствие обработки нуклеазами во время приготовления экстрактов идеально подходит для имитации синергии кэп-поли (A) и других механизмов контроля трансляции, действующих in vivo [56], [63] – [65].Как показано на фиг. 6А, необработанный экстракт сильно зависел от поли (А) хвоста, демонстрируя до 20-кратную стимуляцию трансляции поли (А) хвостом. Важно отметить, что добавление hnRNP-Q2 к экстракту ингибировало трансляцию мРНК с длинными (90–250 нуклеотидами) поли (A) хвостами сильнее (в 3,2–3,6 раза), чем трансляция мРНК с короткими (15–30 нуклеотидов) поли (А) хвосты (в 1,5–1,8 раза), при этом оказывая маргинальное влияние на трансляцию поли (А-) мРНК (1,2-кратное ингибирование). Чтобы гарантировать, что смещение PABP из поли (A) хвоста необходимо для hnRNP-Q2-обеспечиваемого ингибирования трансляции, PABP был изолирован в комплексе PABP-Paip2.В присутствии Paip2 низкоэффективная PABP-независимая трансляция практически нечувствительна к ингибированию hnRNP-Q2 (фиг. 6B). Инактивация PABP также устраняет реакцию трансляции на длину поли (A). Таким образом, ингибирование трансляции hnRNP-Q2 в экстракте, не обработанном нуклеазой, зависит как от PABP, так и от поли (A) хвоста. Чтобы определить, в какой степени эндогенный hnRNP-Q2 ингибирует трансляцию полиаденилированных мРНК, мы попытались истощить необработанный экстракт hnRNP-Q2. Однако нам не удалось достичь существенного иммунного истощения hnRNP-Q2 с использованием антитела 18E4 (неопубликованные данные).Возможно, что hnRNP-Q2 не может взаимодействовать с этим антителом при связывании с мРНК, находящейся в необработанном экстракте. Поскольку в вышеупомянутых анализах использовался экстракт, который не был обработан нуклеазой, представляло интерес проверить влияние hnRNP-Q2 на трансляцию эндогенных мРНК (фигура 6C). HnRNP-Q2 снижал включение 35 S-метионина в необработанный экстракт дозозависимым образом. Однако это ингибирование было относительно небольшим (до 1,4 раза) по сравнению с ингибированием экзогенной трансляции мРНК (рис. 6А).Вероятно, что повторная инициация трансляции в необработанном экстракте менее эффективна, чем в интактных клетках; следовательно, 35 включение S-метионина в первую очередь отражает скорость удлинения полипептидной цепи на предварительно сформированных полисомах. В соответствии с этим представлением, ингибирование повторной инициации трансляции с помощью гиппуристанола [66] лишь незначительно (в 1,8 раза) снижает включение 35 S-метионина (Рисунок 6C).
Рисунок 6. Длина поли (A) хвоста и PABP-зависимое ингибирование трансляции hnRNP-Q2 в экстракте Кребса.
(A) Экстракт Кребса, который не был обработан нуклеазой, был запрограммирован мРНК Cap-Luc (0,2 мкг / мл), несущей поли (A) хвосты указанной длины. Контрольный буфер hnRNP-Q2 (20 мкг / мл) добавляли к реакционным смесям, как указано. (B) Секвестрация PABP с помощью Paip2 делает трансляцию нечувствительной к длине поли (A) хвоста и ингибированию hnRNP-Q2. МРНК Cap-Luc с увеличивающимися поли (A) хвостами транслировали в необработанном экстракте в присутствии Paip2 (15 мкг / мл), как описано для панели A.hnRNP-Q2 (20 мкг / мл) добавляли к реакционным смесям, где указано. Ингибирование трансляции hnRNP-Q2 показано в верхней части панелей. (C) Эндогенное включение [ 35 S] метионина в необработанный экстракт в присутствии указанных концентраций hnRNP-Q2 или 10 мкМ гиппуристанола (Hipp). Инкубация проводилась при 32 ° C в течение 2 часов. Показаны средние значения нерастворимой в трихлоруксусной кислоте радиоактивности в 1-мкл аликвотах образцов из трех анализов со стандартными отклонениями.
https://doi.org/10.1371/journal.pbio.1001564.g006
Нокдаун HnRNP-Q2 стимулирует синтез белка in vivo
Затем мы исследовали, подавляет ли hnRNP-Q2 синтез белка in vivo за счет уменьшения количества hnRNP-Q2 в линии фибробластоподобных клеток мыши L929 с использованием shRNA. Одна shРНК против hnRNP-Q (shRNA1) вызвала значительное подавление hnRNP-Q2 (~ 90%; фиг. 7A, B). Другая shRNA (shRNA2) была менее эффективной в подавлении hnRNP-Q2 (~ 75%).Изменений в уровнях PABP, eIF4GI, eIF4A и eIF4E не обнаружено. Общую скорость трансляции измеряли по включению [ 35 S] метионина / цистеина во вновь синтезированные белки. Клетки, экспрессирующие shRNA1, показали ~ 2-кратное увеличение включения по сравнению с клетками, экспрессирующими нецелевую контрольную shRNA (фигура 7C). Нокдаун hnRNP-Q с помощью shRNA2 вызывал менее сильную стимуляцию трансляции по сравнению с shRNA1 (~ 1,4 раза). Анализ SDS-PAGE вновь синтезированных белков показал, что hnRNP-Q2 ингибирует глобальный синтез белка (рис. 7D).
Рисунок 7. Молчание HnRNP-Q2 стимулирует глобальный синтез белка.
Клетки L929 инфицировали лентивирусами, экспрессирующими нецелевую контрольную shRNA или shRNA (1 и 2) против hnRNP-Q. (A) Цитоплазматические экстракты контрольных клеток и клеток с нокдауном hnRNP-Q, уравновешенные по содержанию белка, подвергали вестерн-блоттингу для определения hnRNP-Q2, PABP, eIF4GI, eIF4A, eIF4E и β-актина, как указано. (B) Количественный анализ полос hnRNP-Q2 на панели A с использованием программного обеспечения NIH ImageJ. Значения были нормализованы по β-актину.Значение в контроле было установлено как 100%. Данные представляют собой средние значения со стандартными отклонениями из трех экспериментов. (C) Синтез белка в контрольных и hnRNP-Q2-нокдаун клетках L929, как анализировали с помощью метионин / цистеинового мечения [ 35 S]. Средние значения для включения 35 S в белки из трех независимых анализов со стандартными отклонениями показаны в процентах от значения в контроле (* p <0,025). (D) Типичные образцы 35 S-меченных белков из панели C.Белки были разделены с помощью SDS 12% PAGE и обнаружены авторадиографией.
https://doi.org/10.1371/journal.pbio.1001564.g007
HnRNP-Q2 антагонизирует активность PABP в miRNA-опосредованном деаденилировании
MiRNA, помимо ингибирования трансляции, опосредуют деаденилирование и распад целевых мРНК [67]. PABP облегчает miRNA-зависимое деаденилирование посредством взаимодействия с комплексом деаденилазы GW182-CAF1 / CCR4 [25], [68]. Мы хотели определить, противодействует ли hnRNP-Q2 этой функции PABP в экстракте Кребса, который точно воспроизводит PABP-зависимое miRNA-опосредованное деаденилирование [25].РНК, несущая шесть сайтов-мишеней let-7a и последовательность поли (A) длиной 98 нуклеотидов (РНК 6xB-3’UTR), однородно меченную 32 P UTP [25], была экстенсивно деаденилирована экстрактами Кребса (рисунки S7 и 8). Полностью деаденилированная (A 0 ) РНК, вероятно, нестабильна, поскольку выглядит как менее заметная полоса по сравнению с входной (A 98 ) РНК. Образование видов РНК A 0 зависело от миРНК let-7a, поскольку она блокировалась добавлением анти-let-7a 2′- O -метилированного олигонуклеотида (2′- O -Me) и также не наблюдается с репортерными мутациями в нуклеотидах, комплементарных «затравочной» последовательности let-7a (РНК 6xBMut-3’UTR) (фигура S7).Чтобы исследовать, как на деаденилирование влияет конкуренция между PABP и hnRNP-Q2, hnRNP-Q2 добавляли к необработанным или обработанным нуклеазой экстрактам Кребса и наблюдали за кинетикой деаденилирования РНК 6xB-3’UTR. Как и ожидалось, экзогенный hnRNP-Q2 ингибировал превращение полноразмерной РНК A 98 в РНК A 0 (фиг. 8A, B). Эффект hnRNP-Q2 на деаденилирование был несколько сильнее в необработанном экстракте, скорее всего, потому, что часть PABP выводится из конкуренции, поскольку она секвестрируется эндогенными мРНК.Чтобы лучше оценить влияние PABP и hnRNP-Q2 на деаденилирование, мы использовали экстракты, лишенные этих белков. В экстракте, обедненном hnRNP-Q2, полноразмерная РНК A 98 исчезла, и РНК A 0 образовалась в течение 2-часового инкубационного периода (фигура 8C). В соответствии с важностью PABP для укорачивания поли (A) хвоста [25], добавление Paip2 почти отменяет деаденилирование. Важно отметить, что добавление hnRNP-Q2 заметно нарушало деаденилирование, поскольку около 30% РНК сохраняли полноразмерный поли (A) хвост через 2 часа инкубации.Это демонстрирует, что конкуренция с hnRNP-Q2 ингибирует опосредованное miRNA деаденилирование. Чтобы определить, влияет ли hnRNP-Q2 также на функцию экзогенного PABP в miRNA-опосредованном деаденилировании, анализ проводили в экстракте Кребса, лишенном как hnRNP-Q2, так и PABP. В этом экстракте значительная часть входящей РНК сохраняла поли (A) хвост в течение реакции (рис. 8D). Однако почти вся РНК деаденилировалась в течение 2 ч после добавления рекомбинантного РАВР.Важно отметить, что добавление hnRNP-Q2 обратно к экстракту, дополненному PABP, заметно снижает скорость деаденилирования. В этих условиях значительная часть РНК (~ 25%) оставалась интактной даже после 3 ч инкубации. Кроме того, слабое деаденилирование РНК в PABP и дважды истощенном экстракте hnRNP-Q2 (что могло быть связано с неполным истощением PABP) предотвращалось добавлением рекомбинантного hnRNP-Q2. Взятые вместе, эти результаты демонстрируют, что hnRNP-Q2 стабилизирует мРНК, противодействуя активности PABP в miRNA-опосредованном деаденилировании.
Фигура 8. HnRNP-Q2 противодействует функции PABP в let-7a miRNA-зависимом деаденилировании.
Кинетика деаденилирования РНК 6xB-3’UTR в необработанных (A) или обработанных нуклеазой (B) экстрактах S10 Krebs. Рекомбинантный hnRNP-Q2 (50 мкг / мл) добавляли к реакционным смесям, где указано. (C) Кинетика деаденилирования РНК 6xB-3’UTR в экстракте Кребса, обедненном hnRNP-Q2. Рекомбинантный hnRNP-Q2 (36 мкг / мл) или GST-Paip2 (16 мкг / мл) были включены в реакционные смеси, где указано. РНК 6xB-3’UTR, обработанная РНКазой H в присутствии олиго (dT), показана на дорожке 1.(D) Кинетика деаденилирования РНК 6xB-3’UTR в дважды истощенном экстракте Кребса hnRNP-Q2 и PABP. Рекомбинантный PABP (6 мкг / мл) и hnRNP-Q2 (36 мкг / мл) включали в реакционные смеси либо по отдельности, либо в комбинации, как указано. Справа указаны положения интактной и деаденилированной РНК. Данные представляют три независимых эксперимента.
https://doi.org/10.1371/journal.pbio.1001564.g008
Истощение HnRNP-Q2 в культивируемых клетках усиливает репрессию, опосредованную miRNA
Затем мы проверили, снижает ли hnRNP-Q2 индуцированную miRNA репрессию in vivo.Контрольные и нокдаунные клетки hnRNP-Q2 L929 трансфицировали репортерами люциферазы (RL) Renilla с шестью сайтами-мишенями миРНК let-7a (6xB) или без них [69]. Репортер светлячка (FL) котрансфицировали для нормализации эффективности трансфекции. В контрольных клетках экспрессия RL-6xB была в ~ 4 раза ниже, чем RL (фиг. 9A, B). Важно отметить, что нокдаун hnRNP-Q2 значительно увеличивал это ингибирование (с 4 до 8,1 раза). Котрансфекция олигонуклеотида anti-let-7a 2′- O -Me, но не контрольного олигонуклеотида anti-miR-122a, резко снизила ингибирование экспрессии RL-6xB, что согласуется с ролью miRNA let-7a в подавлении молчания. репортера RL-6xB (рис. 9А).Мы определили, что количество мРНК RL-6xB в ~ 2,6 раза и в ~ 4 раза ниже, чем мРНК RL в контрольных и нокдаунных клетках hnRNP-Q, соответственно (рис. 9C, вверху; сравните дорожку 2 с 1 и дорожку 8 с 7. ; Рисунок 9D). Эта разница в относительных уровнях RL-6xB может частично объяснить усиленное снижение экспрессии репортера RL-6xB после истощения hnRNP-Q2 (фиг. 9B). Подтверждая зависимость распада мРНК RL-6xB от миРНК let-7a, уровни мРНК RL-6xB были восстановлены путем котрансфекции антителом к let-7a, но не анти-miR-122a, 2′- O -Me олигонуклеотид (фигура 9C).
Рисунок 9. Истощение HnRNP-Q2 усиливает опосредованную miRNA репрессию в культивируемых клетках.
(A) Активность репортеров RL и RL-6xB в контрольных клетках и клетках L929 с нокдауном hnRNP-Q2. Клетки, экспрессирующие контрольную shРНК или hnRNP-Q shRNA2, трансфецировали репортерами RL, имеющими или не имеющими последовательность 6xB, параллельно с репортером люциферазы Firefly (FL). Анти-let-7a или анти-miR-122a (отрицательный контроль) 2′- O -Me олигонуклеотиды котрансфицировали там, где указано.Через два дня после трансфекции измеряли активности RL и FL и определяли их соотношение. (B) Экспрессия RL-6xB относительно RL (которая была установлена как 100% как в контрольных, так и в клетках с нокдауном hnRNP-Q2). (C) Равные количества (10 мкг) РНК из трансфицированных клеток анализировали методом Нозерн-блоттинга с использованием зондов, специфичных для репортеров RL (вверху) и FL (внизу). Полосы были количественно определены с использованием Typhoon PhosphorImager (GE Healthcare). Чтобы скорректировать загрузку и эффективность трансфекции, значения для мРНК RL и RL-6xB были нормализованы к значениям для мРНК FL.Отношение RL-6xB / RL для каждого условия указано под верхней панелью. Обратите внимание, что мРНК RL-6xB мигрирует немного медленнее, чем мРНК RL из-за присутствия последовательности 6xB. (D) Уровни мРНК RL-6xB в контрольных клетках и клетках KD hnRNP-Q были нормализованы к уровням мРНК RL (которые были установлены как 100% для обоих условий). В (A), (B) и (D) значения представляют собой средние значения трех трансфекций со стандартными отклонениями (* p <0,025, ** p <0,001).
https: // doi.org / 10.1371 / journal.pbio.1001564.g009
Обсуждение
В эукариотических клетках ассоциация PABP с поли (A) хвостом стимулирует глобальную трансляцию [12], [17], но также способствует миРНК-зависимому деаденилированию и репрессии целевых мРНК [20], [25], [70]. Paip2 ингибирует эти функции PABP путем диссоциации комплекса PABP-poly (A) [31]. В этом исследовании мы применили УФ-индуцированное сшивание, чтобы охарактеризовать состав поли (A) мРНП в отсутствие PABP. При УФ-облучении белки сшиваются с поли (А), когда они связаны вблизи фотохимически реактивных пуриновых колец [33], [71].Напротив, ионные взаимодействия белков с сахарно-фосфатной основной цепью поли (А) не удовлетворяют требованиям для сшивания. Кроме того, УФ-облучение не вызывает сшивки белок-белок. Таким образом, сшивание, индуцированное УФ-излучением, является надежным методом выявления специфических взаимодействий белок-поли (А). Во всех исследованных клеточных экстрактах PABP проявляется как единственный главный поли (A)-связывающий белок, а Paip2 снижает ассоциацию PABP с поли (A) (рис. 1 и [31], [71]). Когда PABP истощался из RRL, hnRNP-Q2 по умолчанию становился основным связывающим поли (A) белком.PABP и hnRNP-Q2 предположительно являются единственными сильными связующими поли (A), поскольку RRL, обедненный как PABP, так и hnRNP-Q2, не дает серьезных поперечных связей (неопубликованные данные). Насколько нам известно, первое описание цитоплазматического белка, взаимодействующего с поли (A) (p78X), который отличается от PABP, датируется 1981 годом [72]. В то время идентичность этого белка и его функция не были изучены. Наряду с hnRNP-Q2 / Q6, p58 / 59, поперечно сшитый с поли (A) в экстрактах Krebs и HeLa, обедненных PABP (рис. 1B, C). Этот белок (белки) может быть похож или идентичен ядерному поли (A) -ассоциированному белку p60A с еще не идентифицированной функцией [72].Отсутствие p58 / 59 в ретикулоцитах кролика, в которых отсутствуют ядра, способствует этой возможности. Утечка p58 / 59 из ядра могла произойти во время приготовления экстракта, поскольку этих белков особенно много в экстрактах, полученных из чрезмерно поврежденных клеток (неопубликованные данные).
Насколько важна hnRNP-Q2 для трансляции мРНК и метаболизма? Предпочтительное связывание с поли (A) отличает hnRNP-Q2 от основной массы общих РНК-связывающих белков, которые не проявляют специфичности последовательности [73].Более того, hnRNP-Q2 конкурировал с PABP за связывание с поли (A) хвостом мРНК. Ожидается, что эта конкуренция нарушит множество функций PABP в глобальной и мРНК-специфической регуляции синтеза белка. В соответствии с этим предсказанием мы показали, что hnRNP-Q2 ингибирует инициацию трансляции, которая требует комплекса PABP / eIF4G. Кроме того, hnRNP-Q ингибирует опосредованное miRNA деаденилирование и репрессию мРНК, которые стимулируются PABP.
Первостепенная проблема в решении проблемы конкуренции между hnRNP-Q2 и PABP за связывание поли (A) in vivo касается относительного количества этих белков в клетке.Концентрация hnRNP-Q2 в смесях трансляции Krebs и RRL примерно в 1,7 раза выше, чем сообщалось для PABP (240–480 нМ против 140–280 нМ, рис. S3 и [25], [54]). Поскольку сродство hnRNP-Q2 к поли (A) примерно в 6 раз ниже, чем сродство PABP, его молярный избыток над PABP может быть недостаточным для эффективной конкуренции в стандартных физиологических условиях. Однако значительная часть PABP может быть изолирована в комплексы с репрессорными белками, такими как Paip2. Это не только увеличило бы соотношение hnRNP-Q2 / PABP, но также нарушило бы кооперативное связывание PABP, что важно для стабильности комплекса PABP / poly (A).Наконец, цитозольные уровни hnRNP-Q, вероятно, повышены в фазах G2 / M клеточного цикла и в условиях стресса [44], [74]. В результате этих преобразований могут быть выполнены условия для эффективной конкуренции со стороны hnRNP-Q. Наблюдаемые стимулирующие эффекты истощения hnRNP-Q2 на трансляцию как in vitro, так и in vivo указывают на то, что эндогенная концентрация hnRNP-Q2 достаточна для ингибирования трансляции. Хотя анализ белков de novo с помощью SDS-PAGE показывает, что hnRNP-Q2 нацелен на глобальный синтез белка (рис. 7D), он также может по-разному влиять на трансляцию конкретных мРНК.На эту возможность указывает отрицательная регуляция трансляции мРНК RhoA цитоплазматической изоформой hnRNP-Q [36], а также присутствие NSAP1 / hnRNP-Q в комплексе подавления трансляции, который распознает специфический элемент в 3’UTR воспалительного процесса. мРНК (называемые IFN-γ-активированным ингибитором трансляции или GAIT, элементом) [75], [76]. С другой стороны, связывание hnRNP-Q с несколькими элементами IRES стимулирует трансляцию [41] – [45]. Кроме того, hnRNP-Q, возможно, может активировать IRES косвенно, уменьшая конкуренцию со стороны основной массы клеточной мРНК.Наконец, как показано здесь и обсуждается ниже, смещение PABP из поли (A) хвоста с помощью hnRNP-Q2 ослабляет индуцированное miRNA деаденилирование, распад и репрессию специфических мРНК. Т.о., помимо своей функции общего репрессора трансляции, hnRNP-Q может играть разные роли в мРНК-селективном контроле трансляции.
Трансляционно репрессированные мРНК накапливаются в двух цитоплазматических фокусах: процессинговых телах и стрессовых гранулах, которые служат сайтами для деградации или хранения мРНК [77], [78].Возможно, что после связывания hnRNP-Q2 мРНК направляется к цитоплазматическим гранулам. В самом деле, в нейронах SYNCRIP / hnRNP-Q2 локализуется в гранулах мРНК, которые транспортируются по дендритам [79]. Кроме того, в стрессированных клетках hnRNP-Q перемещается в цитоплазматические гранулы, о чем свидетельствует его совместная локализация с белками-маркерами HSP70, GW182 и TIA-1 [74]. В обоих типах гранул трансляция мРНК подавлена [2]. Интересно предположить, что hnRNP-Q2 играет роль в этом ингибировании.
Деаденилирование и последующее снижение уровней мРНК-мишени значительно способствует уменьшению экспрессии генов, индуцированному miRNA [67], [70]. PABP взаимодействует с белками GW182, которые являются важными компонентами miRISC [25]. Это взаимодействие способствует miRNA-зависимому деаденилированию, потенциально за счет приближения поли (A) хвоста к miRISC-ассоциированной деаденилазе CAF1 / CCR4. HnRNP-Q2 заметно нарушает PABP-зависимое let-7a miRNA-опосредованное деаденилирование в экстракте Кребса, наиболее вероятно за счет частичного вытеснения PABP из комплекса поли (A) (фиг. 8).В более физиологическом контексте, в клетках L929 истощение hnRNP-Q2 усиливало зависимую от miRNA деградацию и репрессию мРНК-мишени (фиг.9). Интересно, что на экспрессию репортера RL-6xB более сильное влияние оказывает истощение hnRNP-Q2 на уровне белка, чем на мРНК. Таким образом, вероятно, что hnRNP-Q2 также нацелен на функцию PABP в miRNA-обеспечиваемой репрессии трансляции [20]. Примечательно, что роль hnRNP-Q2 / NSAP1 / SYNCRIP в стабилизации мРНК также подтверждается его присутствием в белковых комплексах, которые стабилизируют мРНК протоонкогена c- fos и c- myc [38], [80] .Таким образом, конкуренция со стороны hnRNP-Q обеспечивает новый механизм, с помощью которого регулируются множественные функции PABP. Контроль функций PABP с помощью hnRNP-Q2 может играть важную роль в различных биологических процессах, таких как развитие, вирусная инфекция и болезнь человека.
Материалы и методы
Белки и антитела
Рекомбинантный PABP, GST-Paip2, eIF4A, eIF4E и eIF4B экспрессировали и очищали, как описано [17], [31], [58], [81]. Нативный eIF4F был очищен от RRL [81].Белки диализовали против буфера А, содержащего 20 мМ трис-HCl, pH 7,5, 100 мМ KCl, 0,1 мМ EDTA, 1 мМ DTT и 10% глицерин. Мышиные моноклональные антитела против hnRNP-Q (клон 18E4) и антитела против FLAG, используемые для приготовления экстрактов Кребса с истощенным и ложным содержанием hnRNP-Q2, соответственно, были от Sigma. Описание антител, используемых для вестерн-блоттинга и иммунопреципитации, см. В соответствующих разделах ниже.
Клонирование, экспрессия и очистка hnRNP-Q2
Вектор экспрессии для мышиного hnRNP-Q2 был сконструирован следующим образом.КДНК, кодирующая hnRNP-Q2 (номер доступа GI: 114145481), была получена с помощью ПЦР с обратной транскрипцией (ОТ-ПЦР) поли (А) + РНК клеток Кребса-2 с использованием набора QIAGEN OneStep RT-PCR. Фрагмент ДНК, кодирующий hnRNP-Q2, был амплифицирован с помощью прямого (GATATACCATGGCTACAGAACATGTTAATGGAAATGGTACTGAAGAGCCTATGGATACTACTTCAGCAG) и обратного (GTGGTGCTCGAGTTGTAACAGGTCAGGACCGGCCTCG6 концевых сайтов и I-концевых Ico5 ‘, разработанных для создания концевых I-концевых сайтов и Ico5′, разработанных для создания концевых Ico. сайту путем введения молчащей мутации (фланкирующие последовательности для целей клонирования и молчащие мутации подчеркнуты).После переваривания Nco I и Xho I фрагмент ДНК, кодирующий hnRNP-Q2, был клонирован в Nco I- Xho I сайтов pET28a (Novagen) для создания вектора для экспрессии hnRNP-Q2 с последовательность из шести His на С-конце (pET28a-hnRNP-Q2-His). Меченный His белок hnRNP-Q2 экспрессировали в Escherichia coli и очищали до очевидной гомогенности хроматографией на агарозе Ni 2+ -нитрилотриуксусной кислоты (QIAGEN) с использованием периодической процедуры.Вкратце, замороженные бактериальные клетки суспендировали в суспензионном буфере (S) (20 мМ HEPES, pH 7,5, 5 мМ 2-меркаптоэтанол и 10% глицерин), содержащем 2 М KCl и полный коктейль ингибиторов протеаз, не содержащий ЭДТА (Roche), и лизировали с помощью обработка ультразвуком. После добавления Triton X-100 (до конечной концентрации 0,1%) остатки клеток удаляли центрифугированием (40000 г , 1 час и 4 ° C). В супернатант добавляли 20 мМ имидазола, pH 7,5 и наносили на агарозную смолу Ni 2+ -NTA, уравновешенную буфером S, содержащим 20 мМ имидазол, 2 М KCl и 0.1% Тритон Х-100. Гранулы промывали сначала тем же буфером, а затем буфером S, содержащим 20 мМ имидазола, 0,1 М KCl и 0,1% Triton X-100. Связанные белки элюировали буфером S, содержащим 250 мМ имидазол и 0,1 М KCl, и подвергали диализу против буфера A.
Препарат мРНК
Плазмиды для транскрипции мРНК Luc с 98-нуклеотидными длинными поли (A) хвостами, T3luc (A) + , T7PVluc (A) + и T7HCVluc (A) + [82], были линеаризованы с помощью Bam HI и транскрибируется с помощью РНК-полимеразы Т3 или Т7, в зависимости от ситуации.Матрицы для транскрипции мРНК Luc с вариабельными поли (A) хвостами были получены с помощью ПЦР с использованием плазмиды T3luc [82], прямого праймера GCTCGAAATTAACCCTCACTAAAGGG и пяти различных обратных праймеров, вместе названных (T) n GGATCCCCCGGGCTGC, где (T) n обозначает олиго (dT) участки из 0, 15, 30, 45 и 90 нуклеотидов (основная последовательность промотора Т3 подчеркнута). После очистки на колонке Chroma Spin-1000 (BD Biosciences) продукты ПЦР транскрибировали с помощью РНК-полимеразы Т3.МРНК Luc с поли (A) хвостом из ~ 250 нуклеотидов получали полиаденилированием мРНК Luc-A 98 с использованием дрожжевой поли (A) полимеразы (USB), как рекомендовано производителем. Кэппирование мРНК осуществляли с использованием системы кэппирования ScriptCap m 7 G (Epicenter). Целостность мРНК проверяли электрофорезом в денатурирующем агарозном геле.
EMSA
Олигонуклеотид синтетической РНК(A 30 , Dharmacon) был 5′-мечен с использованием [γ- 32 P] АТФ и полинуклеотидкиназы.Перед использованием зонд очищали центрифугированием на колонке Chroma Spin-10. Стандартные реакционные смеси связывания (20 мкл) содержали 8 фмоль (~ 40 000 имп / мин) 5′-меченного A 30 , 10 мкл 2-кратного инкубационного буфера (40 мМ HEPES-KOH, pH 7,3, 200 мМ KCl, 4 мМ MgCl 2 , 2 мМ DTT, 0,1% NP40, 10% глицерин и 0,2 мг / мл ацетилированного бычьего сывороточного альбумина) и 2 мкл hnRNP-Q2, разведенных до соответствующих концентраций буфером A. После инкубации при 30 ° C в течение Через 30 мин в образцы добавляли 2 мкл 50% об. / Об. Глицерина и анализировали электрофорезом в 7% неденатурирующем полиакриламидном геле (приготовленном с буфером ТВЕ и 5% об. / Об. Глицерина) при 4 ° C.Полосы визуализировали с помощью авторадиографии. Количество свободной и связанной РНК определяли с помощью Typhoon Phosphorimager (GE Healthcare).
Перевод in vitro
экстрактов Кребса, необработанных или обработанных микрококковой нуклеазой, получали, как описано ранее [34]. Где указано, экстракты, обработанные нуклеазой, были истощены или имитировали истощение hnRNP-Q2 и PABP, как описано ниже. Реакционные смеси (12 мкл) включали экстракты Кребса (50% по объему), соли, важные компоненты трансляции [34], указанные мРНК (0.2 мкг / мл, если не указано в подписях к рисункам), и белки. Для оптимальной трансляции мРНК, содержащих IRES HCV и PV, концентрация KOAc в реакционных смесях была увеличена на 75 мМ. Инкубация проводилась при 32 ° C в течение 1 ч. Люциферазную активность определяли в аликвотах образцов объемом 1 мкл с использованием системы анализа люциферазы (Promega) и биолюминометра Lumat LB 9507 (EG&G Bertold). Приведенные относительные световые единицы (RLU) представляют собой средние значения трех анализов со стандартным отклонением от среднего.
Истощение экстракта Кребса и RRL hnRNP-Q2 и PABP
Перед истощением hnRNP-Q2 обработанный нуклеазой экстракт Кребса был дополнен солями, аминокислотами и системой генерации энергии, как описано ранее [83].Добавленный экстракт Кребса или RRL (Promega) осветляли центрифугированием при 10 000 g в течение 1 мин. Для связывания антитела против hnRNP-Q с шариками 40 мкг (∼20 мкл) анти-hnRNP-Q (18E4) инкубировали с суспензией протеин-G-сефарозы (GE Healthcare; 150 мкл гранулированных шариков на 0,6 мл фосфатного буфера. физиологический раствор [PBS; 137 мМ NaCl, 2,7 мМ KCl, 10 мМ Na 2 HPO 4 и 2 мМ KH 2 PO 4 ]) при 4 ° C в течение 1,5 ч при перемешивании на ротаторе. В контрольной пробирке шарики аналогичным образом инкубировали с 40 мкг антитела против FLAG.После инкубации шарики промывали центрифугированием: один раз PBS, содержащим 1% бычий сывороточный альбумин, и дважды – буфером D (25 мМ HEPES-KOH, pH 7,3, 50 мМ KCl, 75 мМ KOAc и 2 мМ MgCl 2 ). [34]. После заключительного центрифугирования при 2400 g в течение 2 минут гранулы суспендировали в 600 мкл экстракта Кребса (или RRL), уже содержащего необходимые компоненты трансляции. После осторожного перемешивания в течение 1,5 ч шарики осаждали центрифугированием, как описано выше.Супернатанты, которые составляют hnRNP-Q2 и ложно-обедненные экстракты, собирали, снова центрифугировали для удаления любых остаточных шариков и замораживали при -80 ° C небольшими аликвотами. Для истощения PABP добавленный экстракт Кребса или RRL инкубировали с белком GST-Paip2, иммобилизованным на гранулах глутатион-сефарозы [34]. Обедненные экстракты обрабатывали только GST. Для удаления как hnRNP-Q2, так и PABP экстракты инкубировали сначала с анти-hnRNP-Q, а затем с GST-Paip2.Эффективность истощения hnRNP-Q2 и PABP анализировали с помощью вестерн-блоттинга.
УФ сшивание
Поли (A) хвост мРНК кроличьего глобина (1,5 мкг; Gibco BRL, прекращено) удлиняли в общем реакционном объеме 50 мкл с использованием [α- 32 P] АТФ (60 мкКи, 3000 Ки / ммоль, Perkin Elmer ) и дрожжевой поли (A) полимеразы (1500 ЕД, USB) в соответствии с рекомендациями производителя. Инкубация проводилась при 37 ° C в течение 30 мин. После экстракции смесью фенола и хлороформа РНК очищали колоночной хроматографией Chroma Spin-100.Реакционные смеси (15 мкл) содержали 10 мкл RRL (или экстракта Кребса, дополненного необходимыми трансляционными компонентами), меченую РНК (~ 400 000 имп / мин) и другие компоненты, как указано в подписях к рисункам. После инкубации при 30 ° C в течение 10 мин образцы по каплям наносили на стеклянную пластину, покрытую парафильмом, и облучали УФ-светом с длиной волны 254 нм (с использованием бактерицидной лампы мощностью 15 Вт с расстоянием ∼4 см между пластинами). лампа и образцы) в течение 20 мин на льду. Образцы собирали в пробирки, уже содержащие 4 мкл коктейля РНКаз (1 мг / мл РНКазы A и 1000 Ед / мл [∼0.7 мг / мл] нуклеазы S7 [Roche]). Затем добавляли один мкл 200 мМ CaCl 2 и образцы расщепляли при 37 ° C в течение 30 минут. Белки денатурировали добавлением 40 мкл 1,5 × буфера для образца SDS и нагреванием при 95 ° C в течение 5 минут. Образцы анализировали с помощью SDS-PAGE (10% акриламида, 99 ± 1 соотношение акриламид / N, N’-метиленбисакриламид) и авторадиографии при -80 ° C с усиливающим экраном.
Вестерн-блоттинг
Первичные антитела были следующими: мышиные моноклональные антитела против hnRNP-Q (клон 18E4, Sigma), кроличьи поликлональные антитела против PABP [31], кроличьи поликлональные антитела против Paip2 (Sigma), кроличьи поликлональные антитела против YB-1 ( Abcam), кроличьи поликлональные антитела против eIF4GI [84], мышиные моноклональные антитела против eIF4A [85], мышиные моноклональные антитела против eIF4E (BD Biosciences), мышиные моноклональные антитела против рибосомного белка S6 (Santa Cruz) и мышиные моноклональные антитела против eIF4E. антитело против β-актина (Sigma).Белки в образцах разделяли с помощью SDS-PAGE, переносили на нитроцеллюлозную мембрану и детектировали с использованием набора для хемилюминесценции Western Lightning (Perkin-Elmer Life Sciences). Первичные антитела против hnRNP-Q, PABP, eIF4GI, YB-1 и β-актина использовали в разведении 1–2,500, 1–1000, 1–1000, 1–1000 и 1–5000 соответственно. Разведения антител против Paip2, eIF4E и рибосомного белка S6 соответствовали инструкциям производителей. Вторичные конъюгированные с HRP антимышиные или антикроличьи антитела (GE Healthcare), в зависимости от ситуации, использовали в разведении 1-5000.Обычно одну мембрану зондировали, обнажали и снимали перед зондированием другим антителом.
Иммунопреципитация p68
Антитела против hnRNP-Q и анти-PABP, которые использовали для вестерн-блоттинга, также использовали для иммунопреципитации p68. Мышиные моноклональные и кроличьи поликлональные антитела против hsp70 были от Santa Cruz и Calbiochem, соответственно. Для создания 32 P-меченого p68, обедненный PABP RRL подвергали УФ-перекрестной сшивке с мРНК глобина 32 P, меченой поли (A) хвостом, в двадцати аликвотах по 15 мкл.Аликвоты объединяли и обрабатывали РНКазами. После добавления 10% SDS до конечных концентраций 0,2% сшитый RRL разбавляли в 10 раз PBS, содержащим 0,2% NP40. Для иммунопреципитации порции разбавленной реакционной смеси по 0,7 мл инкубировали с шариками, конъюгированными с антителом, протеин-G-сефарозой (20 мкл) при 4 ° C в течение 4 ч при перемешивании на ротаторе. Гранулы трижды промывали 0,2% NP-40, содержащим PBS, и, наконец, только PBS. Связанные белки элюировали нагреванием в буфере для образцов SDS и анализировали с помощью SDS-PAGE и авторадиографии.УФ-сшитый контроль и RRL с обедненными PABP загружали в один и тот же гель для сравнения.
Субклеточное распределение hnRNP-Q2
клеток HeLa S3 выращивали в 15-сантиметровой чашке Петри до ~ 90% слияния. Профиль полисома был получен после центрифугирования свежего клеточного экстракта в градиенте плотности сахарозы 10-50% в соответствии со стандартными методами [86]. Центрифугирование проводили в роторе Beckman SW41Ti при 35000 об / мин в течение 2,5 ч при 4 ° C. Оптическую плотность при 254 нМ непрерывно регистрировали с использованием фракционирующего устройства ISCO (Teledyne ISCO, Lincoln, NE).Аликвоты фракций (30 мкл) анализировали вестерн-блоттингом с использованием антител против hnRNP-Q, PABP, Paip2, YB-1, eIF4A, eIF4E и 40S рибосомного белка S6.
Анализы связывания рибосом
Для исследований связывания с 80S рибосомами 32 P-поли (A) -меченную мРНК глобина (~ 300 000 имп / мин, 6 нг) инкубировали в общем реакционном объеме 30 мкл с обработанным нуклеазой экстрактом Кребса или KCl (60 мМ). -дополненный RRL в присутствии компонентов трансляции и 0,6 мМ циклогексимида [17].Где указано, к реакционным смесям добавляли рекомбинантный hnRNP-Q2 или контрольный буфер. Инкубация проводилась при 32 ° C в течение 15 мин. Реакции останавливали 4-кратным разбавлением ледяным буфером HSB [54], и рибосомные комплексы 80S были разделены центрифугированием в 5-мл 15-30% градиентах сахарозы (ротор Beckman SW55, 54000 об / мин в течение 1 часа, 45 минут). при 4 ° C). Фракции (0,2 мл) собирали с вершины градиентов и анализировали сцинтилляционным счетом. Комплексы 48S образовывались в RRL в присутствии GMPPNP (2 мМ), MgCl 2 (2 мМ) и циклогексимида (0.6 мМ) [17]. Перед добавлением мРНК реакционные смеси предварительно инкубировали при 32 ° C в течение 2 мин. Последующая инкубация с 32 P-поли (A) -меченной мРНК глобина проводилась при 32 ° C в течение 10 минут. Реакции останавливали охлаждением и 4-кратным разбавлением буфером K (20 мМ Трис-HCl, pH 8,0, 2 мМ DTT, 100 мМ ацетат калия), содержащим 5 мМ MgCl 2 [81]. Все реакционные смеси наносили на 11-мл градиенты 10–30% сахарозы, приготовленные с тем же буфером. Центрифугирование проводили в роторе SW41 при 40000 об / мин и 4 ° C в течение 3.5 ч. Фракции (0,36 мл) собирали сверху градиентов. Радиоактивность в каждой фракции определяли и выражали как процент от общего количества восстановленных импульсов. Площадь под пиками (без фона) использовали для количественной оценки связывания рибосом. Профили седиментации очищенных 40S и 60S рибосомных субъединиц в градиентах плотности сахарозы служили для подтверждения положений инициирующих комплексов 80S и 48S.
Анализ химического сшивания
МРНКUncapped Luc (Promega) была 3′-поли (A) удлиненной и радиоактивно помечена в кэпе m 7 G с использованием [α- 32 P] GTP, S-аденозилметионина и гуанилилтрансферазы вируса осповакцины [54] .После очистки и окисления с помощью NaIO 4 РНК, меченная кэпом 32 P, была использована для исследований сшивания в RRL, как описано ранее [17], [54], [87] – [89].
Анализ стабильности мРНК
Для анализа стабильности мРНК in vitro, Cap-Luc-A 98 мРНК, равномерно меченная [α- 32 P] UTP во время транскрипции (1,6 × 10 6 импульсов в минуту, 40 × 10 3 циклов в минуту / нг ), был переведен в экстракты Кребса (ложно-обедненные или обедненные hnRNP-Q2) в общем объеме 100 мкл в стандартных условиях.Аликвоты по 15 мкл реакционной смеси отбирали с 30-минутными интервалами. Тотальную РНК депротеинизировали фенол-хлороформной экстракцией, разделяли на формальдегид-1% агарозном геле и переносили на нейлоновую мембрану (Hybond-N; GE Healthcare). Блоты окрашивали Blot Stain Blue (Sigma) для определения уровней 28S рРНК (контроль загрузки). 32 P-меченный Cap-Luc-A 98 мРНК была обнаружена авторадиографией. Количественные оценки интенсивности полос проводили с использованием программного обеспечения NIH ImageJ.
Масс-спектрометрический анализ
Полосы белка, окрашенного Кумасси, вырезали из геля и обрабатывали трипсином. Триптические пептиды анализировали в Центре инноваций Genome Quebec Innovation Center с использованием системы нано-ВЭЖХ, соединенной с масс-спектрометром 4000 Q TRAP (Applied Biosystems, Foster City, CA). Идентичность пептидов определяли путем поиска в базе данных UniProt (версия 13.8) с ограничением до человека с использованием Mascot (версия 2.1, Matrix Science, Лондон).
HnRNP-Q2 Нокдаун
Мышиная линия L929 подобных фибробластам была приобретена в ATCC.Клетки трансдуцировали двумя shРНК, направленными против человеческого hnRNP-Q, shRNA1 (TRCN0000112054), shRNA2 (TRCN0000112053) и ненаправленной контрольной shRNA (SHC002), используя систему трансдукции лентивируса (Sigma-Aldrich), как рекомендовано производителем. Клетки отбирали пуромицином (2 мкг / мл) в течение 4 дней. Через одну неделю после инфицирования цитоплазматические экстракты были приготовлены и проанализированы на предмет подавления hnRNP-Q2 с помощью вестерн-блоттинга.
Анализ синтеза белка
Контроль и клетки L929 с нокдауном hnRNP-Q2, при ~ 90% конфлюэнтности, промывали DMEM без метионина и инкубировали в DMEM без метионина с добавлением диализованной фетальной бычьей сыворотки (10%; GIBCO), глутамина и [ 35 S] смесь метионина / цистеина для мечения (100 мкКи / мл) при 37 ° C в течение 30 мин.Клетки лизировали в буфере для образцов SDS и определяли включение 35 S в материал, нерастворимый в трихлоруксусной кислоте [34]. Значения включения 35 S были нормированы на количество общего белка в образцах.
Анализ деаденилирования
Для получения репортерной РНК, 350-нуклеотидный фрагмент ДНК pRL-6xB-A 98 , содержащий шесть сайтов-мишеней для человеческой miRNA let-7a [62], амплифицировали с помощью ПЦР с использованием праймеров T7-3’UTR (кодирующих промотор T7 ) и Oligo 3R (-) [25].Полученный продукт ПЦР линеаризовали с использованием сайта рестрикции Age I, расположенного непосредственно после поли (A) хвоста, и использовали в качестве матрицы для транскрипции in vitro. [α- 32 P] UTP-меченая РНК 6xB-3’UTR была синтезирована с использованием набора для транскрипции T7 MaxiScript in vitro (Ambion) и очищена путем пропускания через колонку Mini Quick Spin RNA (Roche) [25], [62] ]. Для анализа деаденилирования части (8 мкл) обработанного нуклеазой и дополненного экстракта Кребса, обедненного либо hnRNP-Q2, либо как hnRNP-Q2, так и PABP, сначала смешивали с указанными количествами рекомбинантного hnRNP-Q2, PABP или Paip2 и затем с 0.1 нг очищенной [α- 32 P] UTP-меченой 6xB-3’UTR РНК в общем объеме 10 мкл. Реакции инкубировали при 30 ° C в течение указанного времени, после чего РНК экстрагировали реагентом TRIzol (Invitrogen) и анализировали электрофорезом в 4,5% полиакриламидно-мочевинном геле. Высушенные гели анализировали с помощью Typhoon PhosphorImager (GE Healthcare).
Анализ репрессии, опосредованной miRNA Let-7a
Почти конфлюэнтные клетки L929, контроль и нокдаун hnRNP-Q2, трансфицировали 100 нг pRL или pRL-6xB и 50 нг pFL в шестилуночных планшетах с использованием Lipofectamine 2000 (Invitrogen) [69].Где указано, 2′- O -Me антисмысловые олигонуклеотиды, комплементарные миРНК Let-7a или miR-122a (Dharmacon), котрансфицировали до конечной концентрации 90 нМ [69]. Клетки разделяли через 24 часа. Активности RL и FL измеряли через 48 часов после трансфекции с использованием набора для анализа на двойную люциферазу (Promega) и определяли их соотношение. Результаты трех оценок представлены как средние значения ± стандартное отклонение. Для анализа уровней мРНК RL и RL-6xB, РНК экстрагировали из запасного набора трансфицированных клеток с помощью TRIzol и подвергали Нозерн-блоттингу, как описано ранее [69], [90].Уровни мРНК RL и RL-6xB нормализовали по контрольной мРНК FL.
Статистический анализ
Статистическая значимость различий между средними значениями оценивалась с использованием парного критерия Стьюдента t с двусторонним распределением. Различия считались достоверными при p <0,05.
Вспомогательная информация
Рисунок S1.
Обозначение p68. (A) Поли (A) взаимодействующие цитоплазматические белки HeLa.Экстракт HeLa S10 был лишен PABP с использованием аффинной матрицы GST-Paip2 и инкубирован с поли (A) сефарозой при 4 ° C в течение 1 ч при перемешивании на ротаторе. Гранулы трижды промывали 0,2 М KCl в буфере B (20 мМ Tris-HCl, pH 7,5, 1 мМ MgCl 2 и 1 мМ DTT). Белки, взаимодействующие с поли (A), последовательно элюировали из шариков 1 M KCl и 2 M LiCl в буфере B, концентрировали и анализировали с помощью SDS-PAGE и окрашивания кумасси синим R-250. Две отдельные полосы (1 и 2) элюата 1 М KCl в области геля при 70 К вырезали и анализировали масс-спектрометрией.(B) Иммунопреципитация p68. RRL, обедненный PABP, подвергали УФ-индуцированному перекрестному связыванию с P-поли (A) хвостом 32 мРНК глобина. Меченые белки иммунопреципитировали кроличьим поликлональным антителом против PABP, мышиным моноклональным антителом против hnRNP-Q (18E4), кроличьим поликлональным антителом против hsp70 или мышиным моноклональным антителом против hsp70, как указано. Осажденные белки разделяли с помощью SDS-PAGE и детектировали с помощью авторадиографии. Также показаны сшивание контрольного RRL и положения маркеров молекулярной массы.(C) Сравнительный вестерн-блоттинг экстрактов RRL, Krebs и HeLa S10 (5 мкл) с использованием антитела 18E4 против hnRNP-Q. Указаны положения hnRNP-Q2 и маркеров молекулярной массы. Сходство молекулярных масс некоторых изоформ hnRNP-Q в клетках HeLa не позволило обеспечить их удовлетворительное разрешение.
https://doi.org/10.1371/journal.pbio.1001564.s001
(TIF)
Рисунок S2.
Субклеточное распределение hnRNP-Q. Цитоплазматический экстракт HeLa фракционировали центрифугированием в градиенте плотности сахарозы.Показаны оптическая плотность (Abs 254 нм) полисом (вверху) и анализ вестерн-блоттинга hnRNP-Q, PABP, Paip2, YB-1, eIF4A, eIF4E и 40S рибосомного белка S6 в аликвотах указанных фракций (внизу). . Появление белка S6 во фракциях 5 и 6 подтверждает идентичность пика 40S.
https://doi.org/10.1371/journal.pbio.1001564.s002
(TIF)
Рисунок S3.
Обилие hnRNP-Q2 в трансляционных смесях. (А) Количественная оценка hnRNP-Q2 в смеси трансляции Кребса.Аликвоты по 5 и 10 мкл полной смеси для трансляции (содержащей 2,5 мкл и 5 мкл экстракта Krebs S10 соответственно) анализировали бок о бок с указанными количествами рекомбинантной hnRNP-Q2. Примерно равные сигналы генерировались 5 мкл смеси для трансляции и 150 нг hnRNP-Q2, устанавливая значение для концентрации hnRNP-Q2 ~ 30 мкг / мл (480 нМ). (B) Количественное определение hnRNP-Q2 в смеси трансляции RRL, выполняемое как в (A). Пять мкл полной смеси для трансляции (эквивалент 3.5 мкл RRL) и 75 нг hnRNP-Q2 генерировали сигналы аналогичной интенсивности, устанавливая значение для концентрации hnRNP-Q2 ~ 15 мкг / мл (240 нМ).
https://doi.org/10.1371/journal.pbio.1001564.s003
(TIF)
Рисунок S4.
Зависимость трансляции от кэп- и поли (A) -хвоста трансляции в обработанных нуклеазой RRL под влиянием концентраций иона калия и мРНК. Указанные концентрации кэпированной или незащищенной мРНК люциферазы светлячка, с поли (A) хвостом или без него (A 98 ), транслировались в RRL, который не был дополнен (A, C) или дополнен (B, D) 60 мМ KCl.Инкубация проводилась при 32 ° C в течение 1 ч. Представлена люциферазная активность в аликвотах образцов объемом 1 мкл и стимуляция трансляции за счет кэппинга мРНК (A, B) и поли (A) хвоста (C и D).
https://doi.org/10.1371/journal.pbio.1001564.s004
(TIF)
Рисунок S5.
Образование комплекса инициации 80S в экстрактах Кребса под действием hnRNP-Q2. Связывание 80S рибосомы с меченной 3′-концом мРНК глобина осуществляли в нормальном (A) или обедненном hnRNP-Q2 (B) экстракте Кребса в присутствии циклогексимида (0.6 мМ). Где указано, к реакционным смесям добавляли HnRNP-Q2 (15 мкг / мл; треугольники) или контрольный буфер (квадраты). Образование инициирующего комплекса 80S анализировали, как описано на фиг. 5A, B.
https://doi.org/10.1371/journal.pbio.1001564.s005
(TIF)
Рисунок S6.
Факторы инициации группы eIF4 снимают опосредованное hnRNP-Q2 ингибирование трансляции. Cap-Luc-A 98 мРНК транслировали в обедненный hnRNP-Q2 экстракт Кребса в присутствии контрольного буфера или рекомбинантного hnRNP-Q2 (15 мкг / мл).Где указано, eIF4F, eIF4A, eIF4E и eIF4B были включены в реакционные смеси при концентрациях 90, 60, 10 и 75 мкг / мл соответственно. Ингибирование трансляции hnRNP-Q2 показано вверху.
https://doi.org/10.1371/journal.pbio.1001564.s006
(TIF)
Рисунок S7.
Let-7a miRNA-зависимое деаденилирование 6xB-3’UTR РНК в экстрактах Кребса. Кинетика деаденилирования РНК 6xB-3’UTR в обработанном нуклеазой экстракте Кребса под действием 10 нМ anti-let-7a 2′- O -Me (Anti-let-7a) или мутаций в нуклеотидах, комплементарных let-7a «начальная» последовательность в 3’UTR (6xBMut-3’UTR).Положения полиаденилированной (A 98 ) и деаденилированной (A 0 ) РНК указаны справа. Данные представляют три независимых эксперимента.
https://doi.org/10.1371/journal.pbio.1001564.s007
(TIF)
Оптогенетический контроль локализации и трансляции мРНК в живых клетках
Буксбаум, А. Р., Хаймович, Г. и Зингер, Р. Х. В нужном месте в нужное время: визуализация и понимание локализации мРНК. Нат. Rev. Mol. Cell Biol. 16 , 95–109 (2015).
CAS Статья Google ученый
Ян Л. и Чен Л. Л. Улучшение инструментария разработки РНК. Наука 358 , 996–997 (2017).
CAS Статья Google ученый
Nelles, D. A. et al. Программируемое отслеживание РНК в живых клетках с помощью CRISPR / Cas9. Cell 165 , 488–496 (2016).
CAS Статья Google ученый
Инголия, Н. Т., Геммагами, С., Ньюман, Дж. Р. и Вайсман, Дж. С. Полногеномный анализ трансляции in vivo с разрешением нуклеотидов с использованием профилирования рибосом. Наука 324 , 218–223 (2009).
CAS Статья Google ученый
Blanchard, S.C., Cooperman, B.S. & Wilson, D.N. Пробная трансляция с помощью низкомолекулярных ингибиторов. Chem. Биол. 17 , 633–645 (2010).
CAS Статья Google ученый
Айзекс, Ф. Дж., Дуайер, Д. Дж. И Коллинз, Дж. Дж. Синтетическая биология РНК. Нат. Biotechnol. 24 , 545–554 (2006).
CAS Статья Google ученый
O’Connell, M. R. et al. Программируемое распознавание и расщепление РНК с помощью CRISPR / Cas9. Nature 516 , 263–266 (2014).
Артикул Google ученый
Лоси А., Гарднер К. Х. и Моглич А. Рецепторы синего света для оптогенетики. Chem. Ред. 118 , 10659–10709 (2018).
CAS Статья Google ученый
Cao, J. et al. Светоиндуцируемая активация трансляции мРНК-мишени в клетках млекопитающих. Chem. Commun. 49 , 8338–8340 (2013).
CAS Статья Google ученый
Lee, S. et al. Обратимая инактивация белков путем оптогенетического захвата в клетках. Нат. Методы 11 , 633–636 (2014).
CAS Статья Google ученый
Ли, С., Ли, К. Х., Ха, Дж. С., Ли, С. Г. и Ким, Т. К. Низкомолекулярные наноузлы как индуцируемые нанозонды для мониторинга динамических молекулярных взаимодействий внутри живых клеток. Angew. Chem. Int. Эд. 50 , 8709–8713 (2011).
CAS Статья Google ученый
Щербакова Д.М., Верхуша В.В. Флуоресцентные белки ближнего инфракрасного диапазона для многоцветной визуализации in vivo. Нат.Методы 10 , 751–754 (2013).
CAS Статья Google ученый
Kleijn, M. et al. Факторы роста нервов и эпидермиса индуцируют синтез белка и активацию eIF2B в клетках PC12. J. Biol. Chem. 273 , 5536–5541 (1998).
CAS Статья Google ученый
Novoa, I. et al. Вызванная стрессом экспрессия генов требует запрограммированного восстановления после репрессии трансляции. EMBO J. 22 , 1180–1187 (2003).
CAS Статья Google ученый
Кедерша, Н., Тисдейл, С., Хикман, Т. и Андерсон, П. Количественная и количественная визуализация стрессовых гранул и обрабатывающих тел млекопитающих в реальном времени. Methods Enzymol. 448 , 521–552 (2008).
CAS Статья Google ученый
Аакалу, Г., Смит, В. Б., Нгуен, Н., Цзян, К. и Шуман, Э. М. Динамическая визуализация локального синтеза белка в нейронах гиппокампа. Нейрон 30 , 489–502 (2001).
CAS Статья Google ученый
Mingle, L.A. et al. Локализация всех семи матричных РНК для комплекса нуклеатора полимеризации актина Arp2 / 3 в выступах фибробластов. J. Cell Sci. 118 , 2425–2433 (2005).
CAS Статья Google ученый
Олейников Ю. и Сингер Р. Х. Визуализация в реальном времени ассоциации ZBP1 с мРНК β-актина во время транскрипции и локализации. Curr. Биол. 13 , 199–207 (2003).
CAS Статья Google ученый
Элискович, С., Шеной, С. М. и Сингер, Р. Х. Визуализация взаимодействия мРНК и белков внутри нейронов. Proc. Natl Acad. Sci. США 114 , E1875 – E1884 (2017).
CAS Статья Google ученый
Шестакова Э.А., Сингер Р. Х. и Кондилис Дж. Физиологическое значение локализации мРНК β-актина в определении полярности и направленной подвижности клеток. Proc. Natl Acad. Sci. США 98 , 7045–7050 (2001).
CAS Статья Google ученый
Katz, Z. B. et al. Компартментализация мРНК β-актина увеличивает стабильность фокальной адгезии и направляет миграцию клеток. Genes Dev. 26 , 1885–1890 (2012).
CAS Статья Google ученый
Sundell, C.L. & Singer, R.H. мРНК актина локализуется в отсутствие синтеза белка. J. Cell Biol. 111 , 2397–2403 (1990).
CAS Статья Google ученый
Парк, Х. Ю., Трчек, Т., Уэллс, А. Л., Чао, Дж. А. и Сингер, Р. Х. Беспристрастный метод анализа для количественной оценки локализации мРНК показывает ее корреляцию с подвижностью клеток. Cell Rep. 1 , 179–184 (2012).
CAS Статья Google ученый
Маттила, П. К. и Лаппалайнен, П. Филоподия: молекулярная архитектура и клеточные функции. Нат. Rev. Mol. Cell Biol. 9 , 446–454 (2008).
CAS Статья Google ученый
Park, H.Y. et al. Визуализация динамики одиночной эндогенной мРНК, меченной в живой мыши. Наука 343 , 422–424 (2014).
CAS Статья Google ученый
Тутуччи, Э. и др. Усовершенствованная система MS2 для точной отчетности о жизненном цикле мРНК. Нат. Методы 15 , 81–89 (2018).
CAS Статья Google ученый
Lionnet, T. et al. Трансгенная мышь для обнаружения in vivo эндогенно меченой мРНК. Нат. Методы 8 , 165–170 (2011).
CAS Статья Google ученый
Shin, Y. et al. Пространственно-временной контроль внутриклеточных фазовых переходов с помощью активируемых светом опто-капель. Cell 168 , 159–171 (2017).
CAS Статья Google ученый
Адамала, К. П., Мартин-Аларкон, Д. А. и Бойден, Е. С. Программируемый РНК-связывающий белок, состоящий из повторов одной модульной единицы. Proc. Natl Acad. Sci. США 113 , E2579 – E2588 (2016).
CAS Статья Google ученый
Kim, J.H. et al. Высокая эффективность расщепления пептида 2А, полученного из тешовируса-1 свиньи, в линиях клеток человека, рыбок данио и мышей. PLoS ONE 6 , e18556 (2011).
CAS Статья Google ученый
Ву, Б., Чао, Дж. А. и Сингер, Р. Х. Флуоресцентная флуоресцентная спектроскопия позволяет получать количественные изображения отдельных мРНК в живых клетках. Biophys. J. 102 , 2936–2944 (2012).
CAS Статья Google ученый
Миямичи, К.и другие. Кортикальные репрезентации обонятельного входа посредством транссинаптического отслеживания. Nature 472 , 191–196 (2011).
CAS Статья Google ученый
Gilbert, L.A. et al. CRISPR-опосредованная модульная РНК-управляемая регуляция транскрипции у эукариот. Cell 154 , 442–451 (2013).
CAS Статья Google ученый
Тайко, Дж., Майер, В. Э. и Хсу, П. Д. Методы оптимизации специфичности редактирования генома CRISPR – Cas9. Мол. Ячейка 63 , 355–370 (2016).
CAS Статья Google ученый
Cong, L. et al. Мультиплексная геномная инженерия с использованием систем CRISPR / Cas. Наука 339 , 819–823 (2013).
CAS Статья Google ученый
Лаукайтис, К. М., Уэбб, Д. Дж., Донай, К. и Хорвиц, А. Ф. Дифференциальная динамика интегрина α5, паксиллина и α-актинина во время образования и разрушения адгезий в мигрирующих клетках. J. Cell Biol. 153 , 1427–1440 (2001).
CAS Статья Google ученый
Gorelik, R. & Gautreau, A. Количественный и беспристрастный анализ направленной персистенции в миграции клеток. Нат.Protoc. 9 , 1931–1943 (2014).
CAS Статья Google ученый
Янг, Х. В., Коллинз, С. Р. и Мейер, Т. Локально возбудимые сигналы Cdc42 управляют клетками во время хемотаксиса. Нат. Cell Biol. 18 , 191–201 (2016).
CAS Статья Google ученый
Буксбаум, А. Р., Ву, Б. и Сингер, Р. Х.Обнаружение одиночной мРНК β-актина в нейронах раскрывает механизм регуляции ее трансляционной способности. Наука 343 , 419–422 (2014).
CAS Статья Google ученый
Хейнс, К. А. и Сильвер, П. А. Синтетическое обращение эпигенетического молчания. J. Biol. Chem. 286 , 27176–27182 (2011).
CAS Статья Google ученый
Ким, Н. Ю., Ли, С. и Хео, В. Д. Оптогенетический контроль локализации и трансляции мРНК в живых клетках. Protoc. Exch . https://doi.org/10.21203/rs.2.20634/v1 (2020).
Контроль трансляции иммунной контрольной точки при раке и его терапевтическое нацеливание
Чен, Д. С. и Меллман, И. Онкология встречается с иммунологией: цикл рак-иммунитет. Иммунитет 39 , 1–10 (2013).
PubMed Статья Google ученый
Casey, S.C. et al. MYC регулирует противоопухолевый иммунный ответ через CD47 и PD-L1. Наука 352 , 227–231 (2016).
CAS PubMed PubMed Central Статья Google ученый
Marzec, M. et al. Онкогенная киназа NPM / ALK индуцирует посредством STAT3 экспрессию иммунодепрессивного белка CD274 (PD-L1, B7-h2). Proc. Natl Acad. Sci. США 105 , 20852–20857 (2008).
CAS PubMed Статья Google ученый
Стюарт, Б. В. и Уайлд, К. Всемирный доклад о раке, 2014 г. (Международное агентство исследований рака, ВОЗ Press, 2014).
Ally, A. et al. Комплексная и интегративная геномная характеристика гепатоцеллюлярной карциномы. Ячейка 169 , 1327 (2017).
Артикул Google ученый
Schlaeger, C. et al. Зависимые от этиологии молекулярные механизмы гепатоканцерогенеза человека. Гепатология 47 , 511–520 (2008).
CAS PubMed Статья Google ученый
Marquardt, J. U. et al. Последовательный анализ транскриптома рака печени человека указывает на позднюю стадию приобретения злокачественных признаков. J. Hepatol. 60 , 346–353 (2014).
CAS PubMed Статья Google ученый
Kaposi-Novak, P. et al. Центральная роль c-Myc во время злокачественного превращения в гепатоканцерогенезе человека. Cancer Res. 69 , 2775–2782 (2009).
CAS PubMed PubMed Central Статья Google ученый
O’Dell, M. R. et al. Мутация Kras (G12D) и p53 вызывает первичную внутрипеченочную холангиокарциному. Cancer Res. 72 , 1557–1567 (2012).
PubMed PubMed Central Статья Google ученый
Saha, S. K. et al. Мутантный IDH ингибирует HNF-4alpha, блокируя дифференцировку гепатоцитов и способствуя развитию рака желчных путей. Nature 513 , 110–114 (2014).
CAS PubMed PubMed Central Статья Google ученый
Гривенников С.И., Гретен Ф. Р. и Карин М. Иммунитет, воспаление и рак. Cell 140 , 883–899 (2010).
CAS PubMed PubMed Central Статья Google ученый
Sato, E. et al. Интраэпителиальные CD8 + лимфоциты, инфильтрирующие опухоль, и высокое соотношение CD8 + / регуляторные Т-клетки связаны с благоприятным прогнозом при раке яичников. Proc. Natl Acad. Sci. США 102 , 18538–18543 (2005).
CAS PubMed Статья Google ученый
Gao, Q. et al. Внутриопухолевый баланс регуляторных и цитотоксических Т-клеток связан с прогнозом гепатоцеллюлярной карциномы после резекции. J. Clin. Онкол. 25 , 2586–2593 (2007).
PubMed Статья Google ученый
Fu, J. L. et al. Увеличение регуляторных Т-клеток коррелирует с нарушением Т-лимфоцитов CD8 и плохой выживаемостью у пациентов с гепатоцеллюлярной карциномой. Гастроэнтерология 132 , 2328–2339 (2007).
PubMed Статья Google ученый
Фридман, В. Х., Пейдж, Ф., Сотес-Фридман, С. и Галон, Дж. Иммунный контекст опухолей человека: влияние на клинический исход. Нат. Rev. Cancer 12 , 298–306 (2012).
CAS PubMed Статья Google ученый
Tumeh, P.C. et al. Блокада PD-1 вызывает ответы, подавляя адаптивную иммунную резистентность. Nature 515 , 568–571 (2014).
CAS PubMed PubMed Central Статья Google ученый
Mellman, I., Coukos, G. & Dranoff, G. Иммунотерапия рака достигает совершеннолетия. Nature 480 , 480–489 (2011).
CAS PubMed PubMed Central Статья Google ученый
Джойс, Дж. А. и Фирон, Д. Т. Исключение Т-клеток, иммунная привилегия и микроокружение опухоли. Наука 348 , 74–80 (2015).
CAS PubMed Статья Google ученый
Kortlever, R.M. et al. Myc взаимодействует с Ras, программируя воспаление и подавление иммунитета. Ячейка 171 , 1301–1315 e1314 (2017).
CAS PubMed PubMed Central Статья Google ученый
Инголия, Н. Т., Геммагами, С., Ньюман, Дж. Р. С. и Вайсман, Дж. С. Полногеномный анализ трансляции in vivo с разрешением нуклеотидов с использованием профилирования рибосом. Наука 324 , 218–223 (2009).
CAS PubMed PubMed Central Статья Google ученый
Butte, M.J., Keir, M.E., Phamduy, T.B., Sharpe, A.H. и Freeman, G.J. Лиганд 1 запрограммированной смерти-1 специфически взаимодействует с костимулирующей молекулой B7-1, подавляя ответы Т-клеток. Иммунитет 27 , 111–122 (2007).
CAS PubMed PubMed Central Статья Google ученый
Herbst, R. S. et al. Прогностические корреляты ответа на анти-PD-L1 антитело MPDL3280A у онкологических больных. Nature 515 , 563–567 (2014).
CAS PubMed PubMed Central Статья Google ученый
Gordon, S.R. et al. Экспрессия PD-1 ассоциированными с опухолью макрофагами подавляет фагоцитоз и иммунитет опухоли. Nature 545 , 495–499 (2017).
CAS PubMed PubMed Central Статья Google ученый
Shalapour, S. et al. Иммунодепрессивные плазматические клетки препятствуют Т-клеточно-зависимой иммуногенной химиотерапии. Nature 521 , 94 – U235 (2015).
CAS PubMed PubMed Central Статья Google ученый
Khan, A.R. et al. В-клетки PD-L1hi являются критическими регуляторами гуморального иммунитета. Нат. Commun. 6 , 5997 (2015).
CAS PubMed Статья Google ученый
Lau, J. et al. Опухоль и клетка-хозяин PD-L1 необходимы для подавления противоопухолевого иммунитета у мышей. Нат. Commun. 8 , 14572 (2017).
Артикул Google ученый
Gollwitzer, E. S. et al. Микробиота легких способствует толерантности к аллергенам у новорожденных через PD-L1. Нат. Med. 20 , 642–647 (2014).
CAS Статья Google ученый
Truitt, M. L. & Ruggero, D. Новые рубежи в трансляционном контроле генома рака. Нат. Ред. Рак 17 , 332 (2017).
CAS PubMed Статья Google ученый
Sendoel, A. et al. Трансляция с нетрадиционных 5′-старт-сайтов приводит к возникновению опухоли. Nature 541 , 494–499 (2017).
Артикул Google ученый
Stern-Ginossar, N. et al. Расшифровка цитомегаловируса человека. Наука 338 , 1088–1093 (2012).
CAS PubMed Статья Google ученый
Calkhoven, C.F., Muller, C. & Leutz, A. Трансляционный контроль экспрессии изоформ C / EBP альфа и C / EBP бета. Gene Dev. 14 , 1920–1932 (2000).
CAS PubMed Google ученый
Palam, L.R., Baird, T.D. и Wek, R.C. Фосфорилирование eIF2 облегчает рибосомный обход ингибирующей восходящей ORF для усиления трансляции CHOP. J. Biol. Chem. 286 , 10939–10949 (2011).
CAS PubMed PubMed Central Статья Google ученый
Starck, S. R. et al. Перевод из 5 ‘нетранслируемой области формирует интегральный стрессовый ответ. Наука 351 , 3867 (2016).
Артикул Google ученый
Sekine, Y. et al. Стрессовые реакции. Мутации в факторе инициации трансляции идентифицируют цель соединения, улучшающего память. Наука 348 , 1027–1030 (2015).
CAS PubMed PubMed Central Статья Google ученый
Сидрауски К., МакГичи А. М. и Инголия Н.T. & Walter, P. Небольшая молекула ISRIB обращает эффекты фосфорилирования eIF2alpha на трансляцию и сборку стрессовых гранул. Элиф 4 , 05033 (2015).
Furic, L. et al. Фосфорилирование eIF4E способствует онкогенезу и связано с прогрессированием рака простаты. Proc. Natl Acad. Sci. США 107 , 14134–14139 (2010).
CAS PubMed Статья Google ученый
Herdy, B. et al. Трансляционный контроль активации фактора транскрипции NF-kappaB и продукции интерферона типа I путем фосфорилирования фактора трансляции eIF4E. Нат. Иммунол. 13 , 543–550 (2012).
CAS PubMed PubMed Central Статья Google ученый
Ueda, T., Watanabe-Fukunaga, R., Fukuyama, H., Nagata, S. & Fukunaga, R. Mnk2 и Mnk1 необходимы для конститутивного и индуцибельного фосфорилирования фактора инициации эукариот 4E, но не для рост или развитие клеток. Мол. Клетка. Биол. 24 , 6539–6549 (2004).
CAS PubMed PubMed Central Статья Google ученый
Reich, S.H. et al. Конструирование на основе структуры пиридон-аминаля eFT508, нацеленного на дисрегулируемую трансляцию, путем селективного ингибирования киназ 1 и 2 (MNK1 / 2), взаимодействующих с митоген-активируемой протеинкиназой. J. Med. Chem. 26 , 3516–3540 (2018).
Артикул Google ученый
Pulko, V. et al. B7-h2, экспрессируемый активированными CD8 Т-клетками, необходим для их выживания. J. Immunol. 187 , 5606–5614 (2011).
CAS PubMed PubMed Central Статья Google ученый
Liu, X. et al. Антитела B7-h2 теряют противоопухолевую активность из-за активации p38 MAPK, что приводит к апоптозу опухолевых CD8 (+) T-клеток. Sci. Отчет 6 , 36722 (2016).
CAS PubMed PubMed Central Статья Google ученый
Betts, M. R. et al. Чувствительная и жизнеспособная идентификация антиген-специфических CD8 + Т-клеток с помощью проточного цитометрического анализа дегрануляции. J. Immunol. Методы 281 , 65–78 (2003).
CAS PubMed Статья Google ученый
Hart, L. S. et al.ER стресс-опосредованная аутофагия способствует Myc-зависимой трансформации и росту опухоли. J. Clin. Инвестировать. 122 , 4621–4634 (2012).
CAS PubMed PubMed Central Статья Google ученый
Kataoka, K. et al. Аберрантная экспрессия PD-L1 из-за нарушения 3′-UTR при множественном раке. Природа 534 , 402 (2016).
CAS PubMed Статья Google ученый
Cerezo, M. et al. Трансляционный контроль иммунного ускользания от опухоли через ось eIF4F-STAT1-PD-L1 в меланоме. Нат. Med. 24 , 1877–1886 (2018).
CAS PubMed Статья Google ученый
D’Cruz, C.M. et al. c-MYC индуцирует онкогенез молочной железы посредством предпочтительного пути, включающего спонтанные мутации Kras2. Нат. Med. 7 , 235–239 (2001).
PubMed Статья Google ученый
Ying, H.Q. et al. Онкогенный крас поддерживает опухоли поджелудочной железы за счет регуляции анаболического метаболизма глюкозы. Cell 149 , 656–670 (2012).
CAS PubMed PubMed Central Статья Google ученый
Шарма П., Ху-Лиескован С., Варго Дж. А. и Рибас А. Первичная, адаптивная и приобретенная устойчивость к иммунотерапии рака. Cell 168 , 707–723 (2017).
CAS PubMed PubMed Central Статья Google ученый
Мацуда Т. и Цепко С. Л. Электропорация и РНК-интерференция в сетчатке грызунов in vivo и in vitro. Proc. Natl Acad. Sci. США 101 , 16–22 (2004).
CAS PubMed Статья Google ученый
Miyazaki, J. et al. Система векторов экспрессии, основанная на промоторе бета-актина курицы, направляет эффективное производство интерлейкина-5. Ген 79 , 269–277 (1989).
CAS PubMed Статья Google ученый
Jackson, E. L. et al. Анализ инициации и прогрессирования опухоли легких с использованием условной экспрессии онкогенных K-ras. Gene Dev. 15 , 3243–3248 (2001).
CAS PubMed Статья Google ученый
Рис, С.и другие. Бицистронный вектор для создания стабильных клеточных линий млекопитающих, который предрасполагает все устойчивые к антибиотикам клетки экспрессировать рекомбинантный белок. Biotechniques 20 , 102–104, 106, 108–110 (1996).
Okada, A., Lansford, R., Weimann, J.M., Fraser, S.E. & McConnell, S.E. Визуализация клеток в развивающейся нервной системе с помощью ретровируса, экспрессирующего модифицированный зеленый флуоресцентный белок. Exp. Neurol. 156 , 394–406 (1999).
CAS PubMed Статья Google ученый
Hsieh, A.C. et al. Трансляционный ландшафт передачи сигналов mTOR управляет инициацией рака и метастазированием. Nature 485 , 55–61 (2012).
CAS PubMed PubMed Central Статья Google ученый
Li, H. & Durbin, R. Быстрое и точное согласование коротких считываний с помощью преобразования Барроуза – Уиллера. Биоинформатика 25 , 1754–1760 (2009).
CAS PubMed PubMed Central Статья Google ученый
Li, B. & Dewey, C. N. RSEM: точное количественное определение транскриптов на основе данных RNA-Seq с референсным геномом или без него. BMC Bioinformatics 12 , 323 (2011).
CAS PubMed PubMed Central Статья Google ученый
Langmead, B. & Salzberg, S. L. Быстрое выравнивание с пропуском чтения с Bowtie 2. Nat. Методы 9 , 357 – U354 (2012).
CAS PubMed PubMed Central Статья Google ученый
Чен, Ю., Лун, А. Т. и Смит, Г. К. От считывания до генов и путей: анализ дифференциальной экспрессии в экспериментах с RNA-Seq с использованием Rsubread и конвейера квази-правдоподобия edgeR. F1000Res. 5 , 1438 (2016).
PubMed PubMed Central Google ученый
Ritchie, M. E. et al. limma обеспечивает анализ дифференциальной экспрессии для секвенирования РНК и исследований на микрочипах. Nucleic Acids Res. 43 , e47 (2015).
PubMed PubMed Central Статья Google ученый
Law, C. W., Chen, Y. S., Shi, W. & Smyth, G. K. voom: прецизионные веса открывают инструменты анализа линейной модели для подсчета считывания RNA-seq. Genome Biol. 15 , R29 (2014).
PubMed PubMed Central Статья Google ученый
Робинсон М. Д. и Ошлак А. Метод масштабной нормализации для анализа дифференциальной экспрессии данных РНК-seq. Genome Biol. 11 , R25 (2010).
PubMed PubMed Central Статья Google ученый
Dobin, A. et al. STAR: сверхбыстрый универсальный выравниватель RNA-seq. Биоинформатика 29 , 15–21 (2013).
CAS PubMed PubMed Central Статья Google ученый
Robinson, J. T. et al. Программа просмотра интегративной геномики. Нат. Biotechnol. 29 , 24–26 (2011).
CAS PubMed PubMed Central Статья Google ученый
Bindea, G. et al. ClueGO: плагин Cytoscape для расшифровки функционально сгруппированных онтологий генов и сетей аннотаций путей. Биоинформатика 25 , 1091–1093 (2009).
CAS PubMed PubMed Central Статья Google ученый
Szklarczyk, D. et al. STRINGv10: сети межбелкового взаимодействия, интегрированные в древо жизни. Nucleic Acids Res. 43 , D447 – D452 (2015).
CAS PubMed Статья Google ученый
Shannon, P. et al. Cytoscape: программная среда для интегрированных моделей сетей биомолекулярного взаимодействия. Genome Res. 13 , 2498–2504 (2003).
CAS PubMed PubMed Central Статья Google ученый
Лю Ф., Сонг Ю. и Лю Д. Трансфекция животных на основе гидродинамики путем системного введения плазмидной ДНК. Gene Ther. 6 , 1258–1266 (1999).
CAS PubMed Статья Google ученый
Козак, М. Анализ удлинения праймеров эукариотических комплексов рибосома-мРНК. Nucleic Acids Res. 26 , 4853–4859 (1998).
CAS PubMed PubMed Central Статья Google ученый
Ran, F. A. et al.Геномная инженерия с использованием системы CRISPR-Cas9. Нат. Protoc. 8 , 2281–2308 (2013).
CAS PubMed PubMed Central Статья Google ученый
Надежный контроль инициации трансляции путем диссоциации фосфорилированных тройных комплексов eIF2α
При инициации трансляции эукариот инициаторная тРНК (Met-tRNA i ) распознает стартовые кодоны AUG в мРНК как часть более крупного комплекса, связанного с 40 S рибосомой.Фактор инициации трансляции 2 эукариот (eIF2) является ключевым фактором, который доставляет Met-тРНК i к рибосомам (Hinnebusch, 2014; Dever et al., 2016). eIF2 представляет собой G-белок и связывает GTP и Met-тРНК и , образуя тройной комплекс (TC) (Schmitt et al., 2010). Вместе с другими факторами инициации TC-связанный рибосомный преинициативный комплекс (PIC) связывается рядом с 5 ’кэпом мРНК и сканирует мРНК обычно с первым кодоном AUG, где взаимодействия антикодона Met-tRNAi / кодона мРНК помогают облегчить инициацию трансляции.Следовательно, TC имеет решающее значение для доставки тРНК i Met к 40 S рибосомам для сканирования распознавания кодонов 5’UTR и AUG (Hinnebusch, 2014; Dever et al., 2016; Llácer et al., 2015). Важно отметить, что у всех изученных эукариот инициация трансляции контролируется путем регулирования активности eIF2, см. Ниже. Met-тРНК i имеет примерно в 10 раз большее сродство к eIF2-GTP, чем eIF2-GDP (Kapp and Lorsch, 2004). Следовательно, гидролиз связанного с eIF2 GTP и высвобождение Pi во время распознавания кодона AUG способствует потере eIF2-GDP из инициирующих рибосом (Algire et al., 2005). Этот eIF2-GDP должен быть преобразован в активную GTP-связанную форму для продолжения активной инициации трансляции. Реактивация eIF2 зависит от eIF2B, многофункционального фактора обмена гуаниновых нуклеотидов (GEF) (Pavitt, 2005).
eIF2B Активность GEF способствует высвобождению GDP из комплексов eIF2-GDP (Panniers et al., 1988; Pavitt et al., 1998), обеспечивая связывание GTP и Met-tRNAi с eIF2. Однако точный механизм действия GEF, опосредованного eIF2B, еще не изучен (Mohammad-Qureshi et al., 2008), см. Рисунок 1A. Одна из высококонсервативных форм контроля трансляции включает передачу сигналов и активацию киназ eIF2, каждая из которых фосфорилирует серин 51 субъединицы eIF2α, например, в периоды клеточного стресса (Pavitt, 2005) и широко известна как интегрированный стресс-ответ (ISR). В ISR фосфорилированная форма eIF2 [eIF2 (αP)] связывает eIF2B непродуктивно, образуя комплекс, ингибируемый GEF, который ограничивает уровни TC (Rowlands et al., 1988; Pavitt et al., 1998) (Figure 1A). Это вызывает общее снижение скорости инициации синтеза белка и в то же время активирует трансляцию ISR-чувствительных мРНК (Young and Wek, 2016).Генетические и биохимические данные показывают, что фосфорилированный eIF2α связывается с регуляторным сайтом на eIF2B, образованным субкомплексом eIF2Bαβδ (Pavitt et al., 1997; Krishnamoorthy et al., 2001; Kashiwagi et al., 2016b). Напротив, функцию GEF обеспечивает eIF2Bε, который взаимодействует с eIF2β и GDP / GTP-связывающей субъединицей eIF2γ (Gomez and Pavitt, 2000; Asano et al., 1999; Alone and Dever, 2006). Субъединица γ eIF2B связывается с eIF2Bε и стимулирует действие GEF (Pavitt et al., 1998; Jennings et al., 2013).
СродствоeIF2 к eIF2B не зависит от гуаниновых нуклеотидов.
( A ) Современная модель активации и ингибирования eIF2 фосфорилированным eIF2. Взаимодействия и действия описаны во введении. ( B ). Обзор анализа равновесного связывания eIF2 / eIF2B. ( C – E ).Аффинность определяли путем смешивания 2 нМ eIF2 с возрастающими концентрациями eIF2B, иммобилизованного на смоле anti-Flag. Смолу Flag осаждали, и оставшийся в каждой фракции супернатанта eIF2 разделяли с помощью SDS-PAGE и подвергали иммуноблоттингу с двумя антителами к субъединице eIF2 (вставка, eIF2α и eIF2γ). Связанную при равновесии фракцию определяли количественным определением: общее (дорожка 0 нМ eIF2B) минус фракция, остающаяся в супернатанте (графики), и использовали для расчета указанных констант диссоциации (нМ ± стандартная ошибка (SE)).Анализы проводили либо без нуклеотида ( C ), либо в присутствии либо 1 мМ GDP ( D ), либо 1 мМ GTP ( E ). ( F ). Мультипликационный вывод рисунка.
https://doi.org/10.7554/eLife.24542.003Второй регулирующий фактор eIF2 – eIF5. eIF5 в первую очередь функционирует как белок, активирующий GTPase (GAP) для eIF2-GTP в преинициативном комплексе 48S (PIC) (Paulin et al., 2001; Algire et al., 2005). Гидролиз GTP внутри eIF2-GTP и последующее высвобождение неорганического фосфата являются ключевыми событиями, которые сигнализируют о распознавании стартовых кодонов AUG посредством инициирования рибосом (Algire et al., 2005). Эти события способствуют высвобождению комплексов eIF2-GDP / eIF5 из инициирующего комплекса, обеспечивая последующее соединение 60S. eIF5 связывает неактивный eIF2-GDP и активный TC с аналогичным сродством (K d = 23 нМ; [Algire et al., 2005]), и мы обнаружили, что eIF5 имеет вторую активность с eIF2-GDP, которая препятствует спонтанному высвобождению GDP из eIF2.Эта функция ингибитора диссоциации GDP (GDI) важна во время ISR, когда eIF2α фосфорилируется, а трансляция ослабляется, поскольку GDI предотвращает независимое высвобождение GDP от eIF2B и обеспечивает жесткий контроль трансляции (Jennings and Pavitt, 2010a, 2010b; Jennings et al., 2016). ). В оптимальных условиях роста клеток, когда доступен свободный eIF2B, мы обнаружили, что eIF2B (в частности, eIF2Bγε) активно вытесняет eIF5 из eIF2-GDP до его действия в качестве GEF (Jennings et al., 2013). Таким образом, комбинированные действия eIF2B и eIF5 регулируют статус нуклеотидов eIF2 (рис. 1A).
GEF обычно функционируют, чтобы активировать своих родственных партнеров-белков G, дестабилизируя связывание Mg 2+ и GDP, вызывая высвобождение GDP с промежуточным продуктом, не содержащим нуклеотидов, который позволяет GTP связываться. Чтобы способствовать обмену нуклеотидов, GEFs обладают более высоким сродством к свободным от нуклеотидов формам своих партнеров G-белков, чем к связанным с нуклеотидами формам (Bos et al., 2007). Например, сродство фактора элонгации трансляции eEF1A / eEF1B составляет 0,125 мкМ, тогда как сродство eEF1B к связанным с нуклеотидами формам eEF1A более чем в 10 раз менее стабильно (K d = 1.5 мкМ) (Gromadski et al., 2007). Для прокариотических EF-Tu и EF-Ts комплекс, свободный от нуклеотидов, в 1000 раз более стабилен (K d = 3 нМ), чем связанные с нуклеотидами формы (K d = 2–6 мкМ) (Gromadski et al. , 2002). eIF2γ структурно консервативен с тРНК-связывающими факторами элонгации eEF1A и EF-Tu, а также с aIF2, обнаруженными у архей, и все они связываются как с GDP / GTP, так и с тРНК (Schmitt et al., 2010). Однако, в отличие от факторов элонгации трансляции, каждый eIF2 / aIF2 имеет по три субъединицы (α-γ), и анализы показывают, что субъединицы eIF2α и β также устанавливают важные контакты с Met-тРНК i внутри TC и модулируют связывание нуклеотидов с eIF2γ. (Хуанг и др., 1997; Naveau et al., 2013; Llácer et al., 2015). eIF2αβ также влияет на взаимодействия с eIF2B и eIF5, как указано выше.
Дополнительная сложность субъединиц eIF2 и eIF2B по сравнению с другими трансляционными G-белками и GEFs подсказывает нам, что eIF2B может не вести себя как типичный GEF. Кроме того, недавние исследования, посвященные взаимодействию EF-Tu с его GEF, EF-Ts, предполагают, что EF-Ts может стимулировать образование TC EF-Tu и может образовывать четвертичный комплекс EF-Tu-GTP / тРНК / EF-Ts (Burnett и другие., 2013, 2014). Эти находки побудили нас исследовать взаимодействие между eIF2B и eIF5 с eIF2-GTP и Met-tRNAi во время активации eIF2 и его контроля с помощью eIF2 (αP). Мы сообщаем о неожиданном открытии того, что на аффинность связывания eIF2B с eIF2 в устойчивом состоянии не влияют гуаниновые нуклеотиды. Мы также сообщаем об антагонистической способности eIF2B, где eIF2B может дестабилизировать TC и конкурировать с tRNAi за eIF2. Мы показываем, что этим новым антагонистическим ролям eIF2B противодействует eIF5, где eIF5 стабилизирует TC, предотвращая дестабилизирующие эффекты eIF2B.Следовательно, eIF5 необходим для устойчивых скоростей образования TC и инициирования перехода. Вместе эти находки подтверждают, что конкуренция между eIF2B и eIF5 за eIF2 продолжается даже после образования TC и что комплексы TC / eIF5 представляют собой надежный продукт обмена нуклеотидов. Наконец, мы демонстрируем, что eIF2B может лучше конкурировать с eIF5 за связывание с TC, образованным с помощью eIF2 (αP), что приводит к повышенной дестабилизации TC. Мы предполагаем, что эти новые взаимодействия eIF2B с TC раскрывают вторую стадию инициации трансляции, регулируемую eIF2 (αP), и что эти события обеспечивают жесткий контроль инициации трансляции.
Пространственный контроль репрессии трансляции и поляризованного роста консервативной киназой NDR Orb6 и РНК-связывающим белком Sts5
[Примечание редакции: ответы автора на первый раунд рецензирования приводятся ниже.]
Благодарим вас за отправку вашей работы под названием «Пространственный контроль репрессии трансляции и поляризованного роста с помощью консервативной киназы NDR Orb6 и РНК-связывающего белка Sts5» на рассмотрение eLife. Вашу статью рецензировали три рецензента, в том числе Аарон Гитлер, а оценку контролировал Дж.Пол Тейлор в качестве редактора-рецензента и Джеймс Мэнли в качестве главного редактора. Наше решение было принято после консультации между рецензентами. Основываясь на этих обсуждениях и отдельных обзорах ниже, мы с сожалением сообщаем вам, что текущая отправка вашей работы не может быть рассмотрена для публикации в eLife, по крайней мере, в ее нынешней форме.
Имейте в виду, что старший редактор, редактор-рецензент и 3 дополнительных рецензента подробно обсудили эту заявку.Описанный механизм – зависимый от фосфорилирования контроль сборки РНК-связывающего белка с Р-тельцами с последующей пространственной и временной регуляцией экспрессии подмножества мРНК – представляет для нас большой интерес. Основным недостатком является недостаточная характеристика взаимодействий мРНК с Sts5, демонстрация пространственного перераспределения этих мРНК на Р-тельца и корреляция с изменениями стабильности мРНК. Эти опасения, в дополнение к тем, которые изложены рецензентами ниже, потребуют более обширной дополнительной работы, чем мы обычно допускаем при пересмотре.Мы просим вернуть исправленные рукописи в течение 2 месяцев. Тем не менее, если вы сможете полностью решить эти проблемы, мы рекомендуем отправить новую рукопись в eLife.
Рецензент № 1:
[…] На мой взгляд, эта рукопись будет интересна читателям eLife и станет важным концептуальным прорывом. Есть только одна область, которую я предлагаю авторам рассмотреть:
Несколько недавних исследований продемонстрировали, что некоторые РНК-связывающие белки, такие как hnRNPA1 и FUS, могут подвергаться разделению жидкость-жидкость in vitro (e.г., Патель и др., Cell 2015; Моллиекс и др., Cell, 2015; Lin et al., Mol Cell 2015). Я предлагаю авторам провести эксперименты in vitro , чтобы увидеть, может ли Sts5 образовывать аналогичные жидкие компартменты и, если да, изменяет ли эти свойства фосфорилирование с помощью Orb6. Одна часть этой истории, которая кажется менее развитой, – это точная роль Orb6 в негативном регулировании агрегации Sts5. Эксперименты in vitro и с определенными компонентами могут помочь лучше определить этот механизм.
БелокSts5 содержит предсказанный неструктурированный N-концевой домен в первых 301 аминокислоте.Мы включили этот вывод в текущую версию статьи (Рисунок 2 – приложение к рисунку 1). Роль неструктурированных доменов в формировании жидких компартментов и цитоплазматических фазовых переходах является биологической проблемой фундаментальной важности, и мы планируем в будущем направить значительные усилия на воссоздание поведения Sts5 in vitro. Однако на данный момент мы считаем, что эти эксперименты выходят за рамки данной статьи, и для их полного и хорошо контролируемого представления потребуется целая дополнительная рукопись.Кроме того, исследования фосфорилирования осложняются существованием других фосфорилированных сайтов в молекуле Sts5, с помощью которых другие киназы могут вносить вклад в его регуляцию (результаты протеомного анализа: Wilson-Grady et al., 2008; Koch et al., 2011; Carpy et al., 2014; Kettenbach et al., 2015), и в настоящее время это может затруднить эксперименты in vitro.
Рецензент № 2:
[…] Эта работа описывает новую роль киназы Orb6 в контроле поляризованного роста независимо от путей Cdc42.Широкий интерес и новизна работы связаны с молекулярной регуляцией сборки и функции тел РНП с помощью консервативной киназы и возможностью видеть эту регуляцию в высокой степени пространственно контролируемой в цитоплазме. Ниже предлагаются эксперименты, которые могут укрепить связи в модели, но в целом я выступаю за публикацию этой рукописи.
1) Доказательства контроля трансляции ограничиваются одним белком (рис. 3). Авторы представляют доказательства, что Orb6 ингибирует Sts5-ассоциацию с P-тельцами.Это, в свою очередь, способствует стабильности и, возможно, трансляции связанных с Sts5 мРНК, кодирующих факторы полярности. Уверенность в универсальности этого механизма была бы повышена, уровни других предполагаемых белков были бы измерены, и если бы Ssp1 был проанализирован более глубоко. Изменяются ли уровни белка Ssp1 у другого мутанта, который не может образовывать Р-тельца? Меняются ли уровни Ssp1 во время клеточного стресса и деления клеток, когда Sts5 явно формирует цитоплазматическую точку?
Далее мы охарактеризовали Efc25, белок, кодируемый связанной с Sts5 мРНК efc25 , уровни которой увеличиваются в клетках sts5Δ и уменьшаются в мутантных клетках orb6 .Мы обнаружили, что уровни белка Efc25 увеличиваются в клетках sts5Δ и уменьшаются в мутантах orb6 , аналогично белку Ssp1 (см. Рисунок 3 – приложение к рисунку 1; рисунок 5 – приложение к рисунку 1E).
Что касается уровней белка Ssp1 у мутантов, которые не могут образовывать Р-тельца, мы протестировали уровни Ssp1 в мутантных клетках pdc1Δ и edc3Δ , которые кодируют компоненты Р-тела, которые регулируют декапирование мРНК. Мы обнаружили, что уровни белка Ssp1 увеличиваются на фоне pdc1Δ и edc3Δ (см. Рис. 3 E и F).
2) Являются ли мРНК, идентифицированные микрочипом, как контролируемые изменяющейся локализацией Sts5, совместимой с моделью пространственного контроля репрессии / деградации / трансляции? Было бы хорошо иметь какое-то другое подтверждение регуляции мРНК помимо микроматриц. Можно ли было бы попробовать одиночную молекулярную РНК FISH и посмотреть, как распределение изменяется от кластерного, как было бы, если бы оно было в больших RNP или распределено по цитозолю? Этот инструмент позволит визуализировать локальные изменения в локализации мишеней мРНК Sts5 в ответ на ингибирование Orb6 или клеточный стресс и поможет различать, потенциально, функции митотической точки Sts5 и Р-телец.
В соответствии с Sts5, контролирующим локализацию мРНК ssp1 , мы обнаружили, что ssp1 флуоресцентных РНК-зондов FISH (Stellaris) совместно локализуются с подмножеством Sts5-GFP и Dcp1-mCherry, содержащих пунту, в голодных по глюкозе клетках (см. Рисунок 3G и Рисунок 3 – приложение к рисунку 1 D). Наши результаты согласуются с экспериментами, опубликованными другими исследователями, например лабораторией Роя Паркера, протокол которой мы адаптировали (Brengues and Parker, 2007).
3) Является ли Sts5 in vivo мишенью для Orb6 в клетках? Результаты подтверждают, что Orb6-зависимое фосфорилирование Sts5 регулирует его ассоциацию с P-тельцами или Rad24.Хотя авт. Представляют результат in vitro киназного анализа , чтобы продемонстрировать, что Orb6 может фосфорилировать Sts5, дальнейшие доказательства того, что Sts5 является прямым субстратом in vivo , будет более убедительным. Это, в свою очередь, позволило бы генерации мутантов, которые не являются фосфорилируемыми, на самом деле продемонстрировать, что фосфорилирование является триггером разборки цитоплазматических точек. Альтернативно, образует ли Sst5 «агрегаты» in vitro и разбираются ли они киназой Orb6?
БелокSts5 содержит неструктурированный N-концевой домен, состоящий из первых 301 аминокислот (рисунок 2 – приложение к рисунку 1).Однако он не рекрутируется в РНП в отсутствие мРНК-связывающего домена. Последовательность Sts5 представляет несколько канонических консенсусных сайтов Orb6 и других консенсусных сайтов Orb6 с более низким сродством, которые также могут нуждаться в модификации для обеспечения максимального ответа. Кроме того, исследования фосфорилирования осложняются существованием других фосфорилированных сайтов другими киназами в разных доменах молекулы Sts5, которые могут способствовать ее регуляции (результаты протеомного анализа (Wilson-Grady et al., 2007; Koch et al., 2011; Carpy et al., 2014; Kettenbach et al., 2015), и это может затруднить эксперименты in vitro. Однако это интересные эксперименты, и в будущем они станут предметом интенсивных усилий лаборатории.
4) Вызывает ли избыточная экспрессия Orb6 неспособность клетки правильно реагировать на стресс?
Нутриционный стресс вызывает рекрутирование Sts5 в RNP и образование P-телец. Таким образом, мы использовали рекрутирование Sts5 в RNPs и формирование P-телец как считывание стресса.Мы наблюдали сверхэкспрессию киназы Orb6 и отслеживали образование Р-телец и рекрутирование Sts5 в РНП при депривации глюкозы. Мы обнаружили, что избыточная экспрессия Orb6 значительно снижает интенсивность Sts5 puncta и P-телец (см. Рисунок 5 – приложение к рисунку 2A).
Рецензент № 3:
В этой рукописи авторы показывают, что летальность NDR киназы Orb6 может быть подавлена потерей функции в РНК-связывающем белке Sts5. Они показывают, что Sts5 фосфорилируется киназой Orb6, которая способствует его ассоциации с 14-3-3; образует агрегаты при голодании или при потере активности Orb6; и связывается с несколькими мРНК, включая мРНК ssp1.Утрата функции Sts5 приводит к увеличению уровней этих и других мРНК, что указывает на то, что Ssp5 способствует деградации мРНК. Они предполагают, что фосфорилирование Orb6 Sts5 способствует его ассоциации 14-3-3, чтобы противодействовать его ассоциации с Р-тельцами, тем самым предотвращая деградацию ключевой мРНК для поляризованного роста. Они также предполагают, что эта регуляция пространственно регулируется с агрегатами Sts5, формирующимися преимущественно на конце нерастущих клеток монополярных клеток; и регулируется во времени, при этом опосредованное Orb6 ингибирование происходит во время митоза и цитокинеза, но снимается при замыкании сократительного кольца.
Идеи, выдвинутые в этом исследовании, интересны и во многом новы. Однако я обнаружил, что некоторые части модели недостаточно хорошо поддерживаются показанными данными с некоторыми несоответствиями. Несколько ключевых моментов в тексте преувеличены по сравнению с приведенными данными, которые в совокупности не могут убедить меня во всем повествовании. Они подробно описаны ниже.
Генетическое взаимодействие sts5 и orb6 (рисунок 1 и рисунок 1 – приложение к рисунку 1):
Авт. Идентифицируют потерю функции sts5 как супрессор летальности мутаций orb6 и показывают некоторые доказательства того, что это подавление отделимо от роли Orb6 в регуляции активности Cdc42.Данные подавления убедительны, но разделение функций менее убедительно. В анализе роста, показанном на рисунке 1 – рисунок в приложении 2A, единичные колонии не видны в более высоких разведениях, и единичный мутантный штамм gef1∆ выглядит очень больным. Исходя из этого анализа, можно утверждать, что двойной мутант gef1∆ orb6-as на самом деле растет лучше, чем одиночный мутант. Это следует уточнить.
Задержка роста штамма gef1Δ , наблюдаемая на фиг. 1 – рисунок в приложении 2А, по сравнению с gef1Δ orb6-as2 в контрольных условиях ДМСО, была вызвана различием в фоновом режиме штаммов.Штамм gef1Δ был отягощен ауксотрофными маркерами ura4Δ и leu1-32 , которые вызывали разницу в росте даже в присутствии добавок, добавленных экзогенно. Мы повторили эти эксперименты с другим штаммом gef1Δ , который не содержит этих мутаций, и не обнаружили разницы в росте между gef1Δ и gef1Δ orb6-as2 в ДМСО. (См. Рисунок 1 – дополнение к рисунку 2A)
Кроме того, если контроль роста orb6-sts5 действительно генетически отделим от пространственного контроля Cdc42, это может предсказывать, что потеря функции Sts5 не корректирует круглую форму мутантов orb6 (при допустимой температуре) из-за активности Cdc42. присутствие на сторонах камеры, т.е.е. sts5-276 orb6-as двойной мутант, обработанный ингибитором PP1 при 25 ° ° C, должен быть круглым. Так ли это?
Действительно, мы проанализировали двойных мутантных клеток sts5Δ orb6-as2 , обработанных ингибитором 1-Na-PP1 при 25 ° C в течение 5 часов, и обнаружили, что они круглые. Мы показываем этот эксперимент в текущей версии статьи (см. Рисунок 1 – приложение к рисунку 1B)
Я не уверен, что точно понимаю, что количественно указано на Рисунке 1 – в приложении к рисунку 1С. На рисунке 1B показано, что 60% клеток orb6-as2 разделены.На рис. 1 – дополнение к рисунку 1 показаны только несептированные клетки с сигналом CRIB на сторонах ячеек, с количественной оценкой, которая составляет 80%. Ясно, что 60 + 80> 100, поэтому я не уверен, как согласовать эти числа. Если количественно определяется только подмножество ячеек, это следует указать.
На рисунке 1 – рисунок в приложении 1C мы количественно оценили степень неправильной локализации CRIB-GFP только в несептирующихся клетках (клетках, которые не содержат перегородки, которая действительно является подмножеством). Мы пояснили это в соответствующей легенде к рисунку 1 – дополнению к рисунку 1C и D.Кроме того, эктопическая локализация CRIB-GFP представляет собой быстрый ответ, который можно наблюдать уже через 15 минут после ингибирования Orb6-as2 с помощью 1-NA-PP1 (Das et al., 2009). Таким образом, экспериментальные условия в эксперименте, изображенном на Рисунке 1 – дополнения к рисункам 1C и 1D, отличаются от условий на Рисунке 1B, на котором мы количественно определили индекс септации после длительного ингибирования Orb6-as2 (2 часа), и доля разделенных клеток была различной в два эксперимента.
Кроме того, на Рисунке 1B авторы указывают стандартное отклонение только от n = 2 экспериментов, которое невозможно вывести.
Как также упоминалось ниже, мы насчитали почти 400 ячеек для каждого условия, что вселяет уверенность в нашем заключении. Рецензент, однако, прав, указывая на проблему стандартного отклонения. В пересмотренной версии мы показываем на рисунке 1A совершенно другой набор из трех экспериментов с использованием штамма sts5Δ вместо аллеля orb4-A9 . Это было сделано в ответ на пункт 6 этого обзора, чтобы упростить использование различных аллелей. Результаты полностью согласуются с предыдущим экспериментом, и 6 экспериментов были выполнены трижды (см. Рисунок 1A).
Формирование точек Sts5, ассоциация с Р-тельцами, деградация мРНК и регуляция с помощью Orb6:
Авт. Показывают, что голодание или ингибирование киназы Orb6 ведет к образованию Sts5 punctae, которые колокализуются с маркером P-телец Dcp1. Потеря функции Sts5 также приводит к снижению количества P-телец после ингибирования Orb6, вовлекая Sts5 в образование P-телец. Однако они не исследуют роль Sts5 в формировании P-тел при голодании, когда структуры P-тел гораздо более заметны.Играет ли Sts5 роль в образовании Р-тел при голодании?
Как упоминалось выше, для изучения роли Sts5 во время пищевого стресса мы провели дополнительные эксперименты, которые не были включены в исходную версию статьи. В текущей версии мы показываем, что потеря sts5 (sts5Δ ) значительно снижает интенсивность P-телец, образующихся после истощения глюкозы. Мы добавили эти эксперименты в дополнительный раздел статьи (Рисунок 5 – приложение к рисунку 2 B-D).
Во время N-голодания похоже, что есть только частичная совместная локализация Sts5 с Dcp1. Об этом следует указать в тексте.
Мы упоминали этот факт в тексте (раздел «Результаты», подраздел «Белки Sts5 рекрутируются в цитоплазматическую точку во время митоза и во время пищевого голодания»).
Что касается уровней мРНК, данные о регуляции уровня мРНК ssp1 убедительны, с эпистазом Sts5 по сравнению с Orb6, что указывает на то, что Orb6 предотвращает опосредованную Sts5 деградацию мРНК Ssp1.Однако ни для одного из других исследованных транскриптов эпистаз не показан. Это необходимо для того, чтобы авторы заявили об общем выводе.
Как упоминалось выше, теперь мы охарактеризуем также efc25 , другую мРНК, которая копуризуется с белком Sts5, уровни которого увеличиваются в клетках sts5Δ и уменьшаются в мутантных клетках orb6 . Подобно белку Ssp1, уровни белка Efc25 увеличиваются в клетках sts5Δ и уменьшаются в мутантах orb6 (см. Рисунок 3 – приложение к рисунку 1; рисунок 5 – приложение к рисунку 1, E).
Связь между образованием агрегатов, Р-тельцами и деградацией мРНК также не ясна. В разделе «Результаты» в подразделе «Киназа Orb6 ингибирует агрегацию Sts5 и локализацию Sts5 в P-тельцах» авторы заявляют, что «киназа Orb6 предотвращает […] локализацию Sts5 в P-тельцах, с эффектом предотвращения деградации специфические мРНК »и в шестом абзаце раздела« Обсуждение », что« Sts5 способствует деградации мРНК, вероятно, путем рекрутирования транскриптов в процессирующие (P) тельца ».Однако наблюдаемый повышенный уровень нескольких мРНК в штамме sts5∆ не коррелирует с совместной локализацией Sts5 с Р-тельцами: последнее проявляется при голодании, тогда как уровни мРНК исследуются в нормальных условиях роста. Сходным образом в rad24∆, где нет очевидной совместной локализации Sts5 с маркерами P-body, авторы утверждают, что уровни Ssp1 снижены (действительно ли это так, также является проблемой – см. Ниже). Из этого следует, что образование агрегатов и регуляция уровня мРНК причинно не связаны.В равной степени возможно, что агрегация является следствием взаимодействия sts5 с / деградацией мРНК или что эти два эффекта или не имеют прямой причинной связи.
В самом деле, наши данные показывают, что в отсутствие Sts5 уровни определенных Sts5-ассоциированных мРНК увеличиваются даже в отсутствие обширного или видимого образования P-телец. Предыдущие открытия S. cerevisiae показывают, что деградация мРНК в определенной степени происходит даже в отсутствие существенного образования Р-телец.Такие процессы, как распад мРНК и РНК-опосредованное молчание генов (Eulalio et al., 2007), а также декапирование мРНК 7 и репрессия трансляции (Decker et al., 2007), функционируют в клетках, не имеющих поддающихся обнаружению микроскопических P-телец, что указывает на то, что даже если P- Компоненты тела играют решающую роль в молчании и распаде мРНК, локализация мРНК в больших Р-тельцах не является необходимой для функционирования. В соответствии с этой идеей, мы обнаружили, что потеря компонентов P-телец Pdc1 и Edc3 приводит к значительному увеличению уровней белка Ssp1, даже в отсутствие стресса, когда P-тела не видны (см. Рис. 3E и F).
Как отмечает автор обзора, наши данные также показывают, что рекрутирование Sts5 в большие RNPs в штамме rad24Δ не вызывает существенного снижения уровней мРНК ssp1 . Однако уровни мРНК Ssp1 снижаются при ингибировании киназы Orb6, что индуцирует как образование Р-телец, так и рекрутирование белка Sts5, в дополнение к другим эффектам, таким как активация Cdc42. Таким образом, наши данные предполагают, что дополнительные, но неизвестные сигналы, зависящие от киназы Orb6, необходимы для индукции деградации мРНК.Мы прояснили наши выводы в текущей версии статьи (например, в разделе «Результаты», подраздел «14-3-3 белок Rad24 отрицательно регулирует рекрутирование Sts5 в цитоплазматические точки»; четвертый абзац раздела «Обсуждение»).
В анализе киназы на фиг. 6А отсутствует отрицательный контроль, чтобы утверждать, что киназа Orb6 фосфорилирует Sts5. Показано, что киназа, связанная с Mob2, фосфорилирует Sts5. Это действительно может быть Orb6, но его следует точно описать в тексте.
Мы изменили текст, чтобы отразить это предостережение (раздел результатов, подраздел «14-3-3 белок Rad24 отрицательно регулирует рекрутирование Sts5 в цитоплазматические точки»).
Анализ киназ и роль 14-3-3:
Эта часть не очень убедительна с текстом и рисунками, демонстрирующими ряд несоответствий и чрезмерных интерпретаций.
Авторы заявляют (раздел результатов, подраздел «14-3-3 белок Rad24 негативно регулирует агрегацию Sts5»), что «уровни мРНК ssp1 постоянно снижаются у мутантов rad24∆», но данные, показанные на рисунке 6E, не подтверждают статистически значительная разница между деформациями wt и rad24∆.Тот факт, что уровни значительно снижены в rad24∆ по сравнению с двойными мутантами sts5∆ или sts5∆ rad24∆, ничего не указывает на возможную роль Rad24, просто то, что Sts5 способствует деградации Ssp1.
Точно так же авторы предлагают далее в разделе «Результаты», что «Orb6 ингибирует… локализацию Sts5 в Р-тельцах, способствуя его взаимодействию с 14-3-3 белком Rad24». Однако данные показывают, что Sts5 формирует агрегаты в клетках rad24∆, но я не вижу доказательств того, что они содержат маркер Dcp1 P-body (см. Рисунок 6C).Авторы действительно пишут, что образование P-тел не сильно индуцируется в rad24∆.
Наш вывод из экспериментов, подробно описанных на фиг. 6C, состоит в том, что одна из ролей киназы Orb6 состоит в том, чтобы регулировать ассоциацию Sts5 с Rad24 и предотвращать рекрутирование Sts5 в RNPs. Наши эксперименты с мутантными клетками rad24Δ действительно показывают, что рекрутирование Sts5 в RNPs происходит независимо от существенного образования Р-телец, поскольку эти клетки не голодают. Мы постарались внести ясность в текущую версию статьи.Мы предполагаем, исходя из наших результатов, и предполагаем, что регуляция белка Sts5 с помощью киназы Orb6 является частью механизма, в конечном итоге регулирующего ассоциацию мРНК с P-тельцами, поскольку мы наблюдаем образование P-телец при ингибировании киназы Orb6. Мы также перефразировали эту часть.
Временное регулирование:
Предлагаемая антикорреляция агрегатов Ssp5 с активностью Orb6 во время деления клеток интересна, но плохо поддерживается. Предыдущая работа показала, что активность Orb6 ингибируется передачей сигналов SIN до закрытия сократительного кольца.Здесь авторы полагаются исключительно на эти опубликованные доказательства, чтобы предложить причинную связь между диссипацией агрегатов Ssp5 во время септации и активностью Orb6. Они должны, по крайней мере, точно определять количество агрегатов с течением времени во время цитокинеза. Для этого лучше всего было бы получить Sts5-3xGFP по времени по сравнению с маркером цитокинеза для определения времени прогрессирования сужения кольца. Они также должны показать, что любое исчезновение агрегатов Sts5 действительно происходит из-за активности Orb6 с использованием либо их аллелей orb6-as или orb6 ts.
Мы количественно определили количество точек Sts5 во время цитокинеза в присутствии или в отсутствие киназы Orb6. В качестве маркера цитокинеза мы использовали Rlc1 в дополнение к калькофлуору. Rlc1, регуляторная легкая цепь миозина II (Le Goff et al., 2000) (Wu et al., 2006), исчезает из места деления, когда актомиозиновое кольцо полностью сокращается и перегородка закрывается (Wei, 2016). Мы обнаружили, что Sts5 puncta присутствует до тех пор, пока Rlc1 присутствует в месте деления (признак того, что закрытие перегородки не завершено), но исчезают в клетках, где Rlc1 покинул место деления, после закрытия перегородки.Указывая на то, что исчезновение точки Sts5 действительно связано с активностью Orb6, мы обнаружили, что точки Sts5 все еще видны в клетках, которые завершили закрытие перегородки (и где Rlc1 покинул место деления), когда активность Orb6 ингибируется. Мы показываем эти эксперименты в текущей версии статьи (см. Рис. 7G и H; рис. 7 – дополнение к рисунку 2).
Локализация Orb6-GFP во время деления клеток, которая, как утверждается, показана на фиг. 7 – приложение 2 к рисунку, не показана. Эта легенда к рисунку также присутствует в нижней части рисунка – приложение 1 к рисунку.
Мы действительно показали локализацию Orb6-GFP на перегородке на рисунке 7 – рисунок в приложении 1A (c), как упоминает рецензент. Рисунок 7 – дополнение к рисунку 2C – это единственная опечатка, за которую мы приносим свои извинения, так как она должна была читать рисунок 7 – приложение к рисунку 1A (c). Мы исправили эту опечатку в тексте.
Фенотип клеточного лизиса клеток sts5∆ должен быть определен количественно.
Мы количественно оценили фенотип лизиса sts5Δ в трех экспериментах и обнаружили, что в наших экспериментальных условиях 43% разделяющих клеток sts5Δ лизируются при 36 ° ° C по сравнению с 8% разделяющими контрольными клетками.Мы добавили эту информацию в соответствующую легенду (см. Рис. 7 – приложение к рисунку 2).
Пространственное регулирование:
В целом, пространственная регуляция этого контроля стабилизации мРНК не очень убедительна. Асимметрию агрегатов Sts5 увидеть довольно сложно. Это хорошо задокументировано в клетках tea1∆, но в меньшей степени в клетках wt. Не могли бы авторы предоставить больше примеров, чтобы показать асимметричную локализацию Sts5 в монополярных клетках wt? Авторы предполагают существование градиента фосфорилирования Sts5.Это интересная идея с концептуальным сходством с градиентами MEX-5 у C.elegans, которые способствуют растворению P-гранул, но возможно ли это на самом деле в небольшом масштабе дрожжевой клетки? Скорости диффузии в цитозоле, если Sts5 диффундирует беспрепятственно, делают это маловероятным, но это может быть ограничено за счет образования агрегатов. Разобраться в этом вопросе поможет изучение подвижности агрегатов Sts5.
Действительно, чтобы подтвердить наблюдение асимметричной точки Sts5 в небольших монополярных клетках, впервые наблюдаемой в клетках дикого типа, мы использовали клетки tea1Δ , поскольку они на 9 длиннее, остаются монополярными и локализуют Orb6-GFP только на одном конце.Мы добавили больше примеров небольших клеток дикого типа, которые отражают среднюю репрезентативную локализацию в условиях активного роста клеток (см. Рисунок 7 – приложение к рисунку 1A, D, E и F). Чтобы избежать путаницы или преувеличений, мы убрали слово «градиент», которое изначально предлагалось в ходе обсуждения.
Я также не уверен, что Sts5 играет прямую роль в регуляции биполярного роста. Поскольку мутантные клетки sts5∆ длиннее при всех исследованных плотностях, увеличенная доля биполярных клеток может быть просто результатом большей длины клеток.Хорошо известно, что делящиеся дрожжевые клетки обладают зависимой от размера клетки регуляцией биполярности. Авторы вместо этого, кажется, утверждают, что увеличение длины является следствием повышенной биполярности, но я не вижу доказательств этому. Для клеток одинаковой начальной длины растут ли мутантные клетки sts5∆ быстрее? Являются ли клетки одинаковой длины более биполярными, если sts5∆?
В ответ на комментарии этого рецензента мы выполнили две дополнительные серии экспериментов. Мы показываем, что клеток sts5Δ демонстрируют повышенную биполярность даже во время экспоненциального роста клеток, когда клеток sts5Δ имеют одинаковый размер (средняя длина клетки при делении; см. Фигуру 8C).Как предложил этот рецензент, мы измерили процент биполярных клеток в клетках аналогичной длины (менее 10 микрометров; от 10 до 12; более 12) и обнаружили, что sts5Δ клеток демонстрируют повышенный процент биполярных клеток по сравнению с контроли (см. фигуру 8 – приложение к рисунку 1, A), даже если клеток sts5Δ демонстрируют ту же скорость роста, что и контрольные sts5 + клеток (фигура 8 – приложение к рисунку 1B).
Номенклатура аллелей и температурная чувствительность:
Номенклатура используемых аллелей сбивает с толку.В тексте должно быть четко указано, что orb4 и sts5 – это два названия одного и того же гена (и, насколько это возможно, используйте одно имя гена во всей рукописи), и что используемые аллели являются обоими ранними стоп-кодонами. Авторы также используют делецию гена, но не очень ясно, какой штамм используется в каком эксперименте и почему использовались разные штаммы. Например, во втором абзаце раздела «Результаты» авторы говорят, что они «исследовали, подавляет ли делеция sts5 также потерю полярности, наблюдаемую при ингибировании киназы Orb6», но на приведенном рисунке (Рисунок 1 – рисунок в приложении 1B-C) указано использование аллель orb4-A9.
Мы переделали эксперименты на рисунке 1A, используя аллель sts5Δ , и получили идентичные результаты с аллелем orb4-A9 . Помимо экспериментов по подавлению роста, показанных на рисунке 1E (проведенных некоторое время назад с аллелем sts5-276 нашими сотрудниками из Хиросимского университета) и одного контрольного эксперимента с CRIB-GFP (который был частью первоначального скрининга с orb4-A9 Аллель ; Рисунок 1 – рисунок приложения 1C, D), все остальные эксперименты проводили с использованием аллеля sts5Δ .Как отмечает обозреватель, все это ранние стоп-кодоны, и мы никогда не обнаружили, что соответствующие мутанты ведут себя иначе, чем мутантные клетки с делецией sts5Δ . Мы упомянули этот аспект в легенде к рисунку 1 – добавлению к рисунку 1.
В целом, я не понимаю использования различных «разрешающих», «полупермиссивных» и «ограничительных» температур для изучения фенотипа потери функции sts5. Каким образом используются условные аллели, если это ранние стоп-кодоны или делеции? Об этом следует четко заявить и обсудить.Например, показано, что повышенные уровни Ssp1-HA у мутантов sts5 происходят только при 36 ° C, но в тексте указано, что повышенные уровни «преувеличены» при 36 ° C (раздел результатов, подраздел «Потеря Sts5 приводит к повышенные уровни мРНК, участвующих в контроле роста и активации биполярного роста »), что означает, что это также имеет место при более низкой температуре.
В текущей версии статьи для ясности мы решили не использовать «ограничительные» или «разрешающие» определения, а просто указать температуру, при которой проводятся эксперименты.Использование «ограничительной» или «разрешающей» температуры относится к условным, чувствительным к температуре фенотипам и, таким образом, является функциональным определением. Действительно, sts5Δ демонстрирует морфологический фенотип, чувствительный к температуре: он имеет цилиндрическую форму (хотя и с увеличением степени роста клеток на новом конце) при 25 ° C, в то время как он увеличен и округлен при 36 ° ° C. Эти фенотипы, вероятно, отражают пониженную способность клеток sts5Δ адаптироваться к более высоким температурам.Мы изменили формулировку седьмого абзаца раздела «Результаты».
[Примечание редакции: ответы автора на повторную рецензию приводятся ниже.]
Благодарим вас за отправку вашей статьи «Пространственный контроль репрессии трансляции и поляризованного роста с помощью консервативной киназы NDR Orb6 и РНК-связывающего белка Sts5» на рассмотрение eLife. Ваша статья была рецензирована тремя рецензентами, в том числе Аароном Д. Гитлером (рецензент №1), а оценка проводилась под наблюдением редактора-рецензента и Джеймса Мэнли в качестве старшего редактора.
Рецензенты обсудили рецензии друг с другом, и редактор-рецензент подготовил это решение, чтобы помочь вам подготовить исправленную заявку. Мы надеемся, что вы сможете отправить исправленную версию в течение двух месяцев, поэтому сначала сообщите нам, если у вас возникнут какие-либо вопросы.
Резюме:
Эта рукопись представляет собой новое понимание связи между регуляцией мРНК и поляризованным ростом S. pombe. Авторы представляют генетические и биохимические доказательства, указывающие, что киназа NDR Orb6 негативно регулирует РНК-связывающий белок Sts5.Активность Orb6 изменяет взаимодействие Sts5 с 14-3-3, регулируя способность Sts5 собираться в цитоплазматические Р-тельца, которые секвестрируют и влияют на стабильность мРНК-мишеней Sts5. Рецензенты пришли к консенсусу в отношении того, что дополнительные данные и анализ, представленные в пересмотренной рукописи, решают большинство проблем, возникших в результате первоначального обзора, но остаются некоторые вопросы, которые необходимо решить.
Существенные изменения:
1. Хорошим дополнением является эксперимент FISH, показывающий совместную локализацию мРНК ssp1 с гранулами Sts5 (и Dcp1).Однако, помимо демонстрации репрезентативных изображений, необходимо количественно оценить степень колокализации.
Мы количественно определили совместную локализацию мРНК ssp1 и гранул Sts5 (и Dcp1) и добавили эти данные на рис. 4 – приложение к рисунку 3B и C. Мы использовали подключаемый модуль ImageJ JACoP (Just Another Colocalization Plugin) для выполнения объекта количественная оценка на основе расстояния между центрами масс точек (Bolte and Cordelieres, 2006). Мы измерили, соответственно, долю точки ssp1 , совместно локализующихся с Sts5-3xGFP или Dcp1-mCh (рис. 4 – приложение к рисунку 3B), и долю точки Sts5-3xGFP или Dcp1-mCh, колокализирующуюся с пунктами ssp1 (рис. 4 – приложение к рисунку 3С) в трех независимых экспериментах в присутствии или в отсутствие глюкозы.
2. Точно так же анализ уровней белка Efc25 является хорошим дополнением, хотя его необходимо количественно оценить. Эксперимент с эпистазом, чтобы проверить, отменяет ли делеция sts5 снижение уровней Efc25, наблюдаемое у мутанта orb6 (как показано для Ssp1), дополнительно подтвердит вывод.
Мы провели дополнительные эксперименты и показали, что уровни Efc25 снижаются у чувствительных к ингибитору мутантов orb6-as2 (Рисунок 6 – рисунок в приложении 1E). Мы также обнаружили, что делеция sts5 отменяет снижение уровней Efc25 у мутантов orb6-as2 (Рисунок 6 – рисунок в приложении 1E).Мы количественно оценили результаты трех независимых экспериментов на рисунке 6 – приложение к рисунку 1E, правая панель.
3. На рисунке 6E статистические сравнения должны выполняться между rad24∆ и wt, а не между rad24∆ и sts5∆.
Мы изменили рисунок 7E (рисунок 8E в текущей рукописи) соответствующим образом, чтобы отразить идею эксперимента, что нет существенной разницы в уровнях мРНК ssp1 между rad24∆ и wt.Значения P определяли с использованием дисперсионного анализа (ANOVA) с пакетом статистики SPSS 22.0, с последующим апостериорным тестом Games-Howell. Мы также удалили двойной мутант sts5∆ rad24∆ , потому что, как заявили рецензенты, он не добавлял смысла рисунка.
https://doi.org/10.7554/eLife.14216.030Контроль трансляции экспрессии гена энтеровируса A71 | Journal of Biomedical Science
Schmidt NJ, Lennette EH, Ho HH. По-видимому, новый энтеровирус, выделенный от пациентов с заболеванием центральной нервной системы.J Infect Dis. 1974. 129 (3): 304–9.
CAS PubMed Статья PubMed Central Google ученый
Хо М., Чен Э.Р., Хсу К.Х., Тву С.Дж., Чен К.Т., Цай С.Ф. и др. Эпидемия энтеровирусной 71 инфекции на Тайване. Тайваньская рабочая группа по эпидемии энтеровируса. N Engl J Med. 1999. 341 (13): 929–35.
CAS PubMed Статья PubMed Central Google ученый
Ooi MH, Wong SC, Lewthwaite P, Cardosa MJ, Solomon T. Клинические особенности, диагностика и лечение энтеровируса 71. Lancet Neurol. 2010. 9 (11): 1097–105.
PubMed Статья PubMed Central Google ученый
Yuan J, Shen L, Wu J, Zou X, Gu J, Chen J, et al. Белки энтеровируса А71: структура и функции. Front Microbiol. 2018; 9: 286.
PubMed PubMed Central Статья Google ученый
Лин Джи, Чен Т.С., Вен К.Ф., Чанг СК, Чен Л.Л., Ши С.Р. Белки вируса и хозяина, участвующие в жизненном цикле пикорнавирусов. J Biomed Sci. 2009; 16: 103.
PubMed PubMed Central Статья CAS Google ученый
Бартон Д.Дж., О’Доннелл Б.Дж., Фланеган Д.Б. 5′-клеверный лист в РНК полиовируса является цис-действующим элементом репликации, необходимым для синтеза отрицательной цепи. EMBO J. 2001; 20 (6): 1439–48.
CAS PubMed PubMed Central Статья Google ученый
Thompson SR, Sarnow P. Энтеровирус 71 содержит элемент IRES типа I, который функционирует при расщеплении эукариотического фактора инициации eIF4G. Вирусология. 2003. 315 (1): 259–66.
CAS PubMed Статья PubMed Central Google ученый
Kok CC, Phuektes P, Bek E, McMinn PC. Модификация нетранслируемых областей человеческого энтеровируса 71 ослабляет рост клеточно-специфическим образом. J Virol. 2012. 86 (1): 542–52.
CAS PubMed PubMed Central Статья Google ученый
Lloyd RE, Jense HG, Ehrenfeld E. Ограничение трансляции кэпированной мРНК in vitro в качестве модели индуцированного полиовирусом ингибирования синтеза белка клетки-хозяина: связь с расщеплением p220. J Virol. 1987. 61 (8): 2480–8.
CAS PubMed PubMed Central Статья Google ученый
Sommergruber W., Ahorn H, Klump H, Seipelt J, Zoephel A, Fessl F, et al. 2А протеиназы коксаки- и риновирусов расщепляют пептиды, полученные из гамма-белка eIF-4 через общий мотив узнавания.Вирусология. 1994; 198 (2): 741–5.
CAS PubMed Статья PubMed Central Google ученый
Гради А., Свиткин Ю.В., Иматака Х., Соненберг Н. Протеолиз человеческого фактора инициации трансляции эукариот eIF4GII, но не eIF4GI, совпадает с отключением синтеза белка хозяина после инфекции полиовируса. Proc Natl Acad Sci U S. A. 1998; 95 (19): 11089–94.
CAS PubMed PubMed Central Статья Google ученый
Lamphear BJ, Yan R, Yang F, Waters D, Liebig HD, Klump H и др. Картирование сайта расщепления в факторе инициации синтеза белка eIF-4 гамма протеаз 2A из вируса Коксаки и риновируса человека. J Biol Chem. 1993. 268 (26): 19200–3.
CAS PubMed PubMed Central Google ученый
Haghighat A, Svitkin Y, Novoa I, Kuechler E, Skern T., Sonenberg N. Комплекс eIF4G-eIF4E является мишенью для прямого расщепления протеиназой риновируса 2A.J Virol. 1996. 70 (12): 8444–50.
CAS PubMed PubMed Central Статья Google ученый
Krausslich HG, Nicklin MJ, Toyoda H, Etchison D, Wimmer E. Протеиназа 2A полиовируса индуцирует расщепление полипептида p220 фактора инициации эукариот 4F. J Virol. 1987. 61 (9): 2711–8.
CAS PubMed PubMed Central Статья Google ученый
Ли В., Росс-Смит Н., Гордый К. Г., Белшем Дж. Дж. Расщепление фактора инициации трансляции 4AI (eIF4AI), но не eIF4AII, протеазой 3C вируса ящура: идентификация сайта расщепления eIF4AI. FEBS Lett. 2001; 507 (1): 1–5.
CAS PubMed Статья PubMed Central Google ученый
de Breyne S, Bonderoff JM, Chumakov KM, Lloyd RE, Hellen CU. Расщепление фактора инициации эукариот eIF5B протеазами энтеровируса 3C.Вирусология. 2008. 378 (1): 118–22.
PubMed PubMed Central Статья CAS Google ученый
Kuyumcu-Martinez NM, Joachims M, Lloyd RE. Эффективное расщепление связанного с рибосомами поли (A) -связывающего белка протеазой 3C энтеровируса. J Virol. 2002. 76 (5): 2062–74.
CAS PubMed PubMed Central Статья Google ученый
Куюмку-Мартинес Н.М., Ван Иден М.Э., Юнан П., Ллойд РЭ.Расщепление поли (А) -связывающего белка протеазой полиовируса 3С ингибирует трансляцию клетки-хозяина: новый механизм остановки трансляции хозяина. Mol Cell Biol. 2004. 24 (4): 1779–90.
CAS PubMed PubMed Central Статья Google ученый
Joachims M, Van Breugel PC, Lloyd RE. Расщепление поли (A) -связывающего белка протеазами энтеровируса одновременно с ингибированием трансляции in vitro. J Virol. 1999. 73 (1): 718–27.
CAS PubMed PubMed Central Статья Google ученый
Wu S, Wang Y, Lin L, Si X, Wang T, Zhong X и др. Протеаза 2A индуцирует образование стрессовых гранул при инфицировании вирусом Коксаки B3 и энтеровирусом 71. Вирол Дж. 2014; 11: 192.
PubMed PubMed Central Статья CAS Google ученый
Кедерша Н., Чо М.Р., Ли В., Яконо П.В., Чен С., Гилкс Н. и др.Динамическое перемещение TIA-1 сопровождает рекрутирование мРНК на стрессовые гранулы млекопитающих. J Cell Biol. 2000. 151 (6): 1257–68.
CAS PubMed PubMed Central Статья Google ученый
Jheng JR, Lau KS, Tang WF, Wu MS, Horng JT. Стресс эндоплазматического ретикулума индуцируется и модулируется энтеровирусом 71. Cell Microbiol. 2010. 12 (6): 796–813.
CAS PubMed Статья PubMed Central Google ученый
Чанг Й.Х., Лау К.С., Куо Р.Л., Хорнг Дж.Т. Связывающий домен дцРНК PKR протеолитически высвобождается энтеровирусом A71 для облегчения репликации вируса. Front Cell Infect Microbiol. 2017; 7: 284.
PubMed PubMed Central Статья CAS Google ученый
White JP, Reineke LC, Lloyd RE. При заражении полиовирус переключается в режим трансляции, не зависящий от eIF2. J Virol. 2011; 85 (17): 8884–93.
CAS PubMed PubMed Central Статья Google ученый
Du Z, Ульянов Н.Б., Ю. Дж., Андино Р., Джеймс ТЛ. Структуры ЯМР петли B РНК из домена стебель-петля IV внутреннего сайта рибосомы энтеровируса: одиночная замена C на U резко изменяет форму и гибкость РНК. Биохимия. 2004. 43 (19): 5757–71.
CAS PubMed Статья PubMed Central Google ученый
de Breyne S, Yu Y, Unbehaun A, Pestova TV, Hellen CU. Прямое функциональное взаимодействие фактора инициации eIF4G с внутренними сайтами входа в рибосомы 1 типа.Proc Natl Acad Sci U S. A. 2009; 106 (23): 9197–202.
PubMed PubMed Central Статья Google ученый
Николсон Р., Пеллетье Дж., Ле С.Ю., Соненберг Н. Структурный и функциональный анализ посадочной площадки рибосомы полиовируса типа 2: исследования трансляции in vivo. J Virol. 1991. 65 (11): 5886–94.
CAS PubMed PubMed Central Статья Google ученый
Суини Т.Р., Абаева И.С., Пестова Т.В., Эллен Кр. Механизм инициации трансляции на IRES пикорнавируса 1 типа. EMBO J. 2014; 33 (1): 76–92.
CAS PubMed Статья PubMed Central Google ученый
Лукавский П.Ж. Структура и функция доменов IRES вируса гепатита C. Virus Res. 2009. 139 (2): 166–71.
CAS PubMed PubMed Central Статья Google ученый
Holcik M, Sonenberg N. Трансляционный контроль при стрессе и апоптозе. Nat Rev Mol Cell Biol. 2005. 6 (4): 318–27.
CAS PubMed Статья PubMed Central Google ученый
Левенгуд Дж. Д., Толберт М., Ли М. Л., Толберт Б. С.. Для трансляции и репликации энтеровируса 71 требуется высокоаффинное взаимодействие hnRNP A1 со структурными элементами консервативной РНК. RNA Biol. 2013. 10 (7): 1136–45.
CAS PubMed PubMed Central Статья Google ученый
Lin JY, Shih SR, Pan M, Li C, Lue CF, Stollar V и др. hnRNP A1 взаимодействует с 5′-нетранслируемыми областями энтеровируса 71 и РНК вируса Синдбис и требуется для репликации вируса. J Virol. 2009. 83 (12): 6106–14.
CAS PubMed PubMed Central Статья Google ученый
Tolbert M, Morgan CE, Pollum M, Crespo-Hernandez CE, Li ML, Brewer G, et al. HnRNP A1 изменяет структуру консервативного домена IRES энтеровируса для стимуляции трансляции вируса.J Mol Biol. 2017; 429 (19): 2841–58.
CAS PubMed PubMed Central Статья Google ученый
Hellen CU, Witherell GW, Schmid M, Shin SH, Pestova TV, Gil A, et al. Цитоплазматический белок 57 кДа, который необходим для трансляции РНК пикорнавируса посредством внутреннего входа в рибосомы, идентичен ядерному белку, связывающему пиримидиновый тракт. Proc Natl Acad Sci U S. A. 1993. 90 (16): 7642–6.
CAS PubMed PubMed Central Статья Google ученый
Xi J, Ye F, Wang G, Han W, Wei Z, Yin B и др. Белок, связывающийся с полипиримидиновым трактом, регулирует трансляцию энтеровируса 71 посредством взаимодействия с внутренним сайтом входа в рибосомы. Virol Sin. 2019; 34 (1): 66–77.
PubMed PubMed Central Статья Google ученый
Кафасла П., Моргнер Н., Робинсон К.В., Джексон Р.Дж. Белок, связывающийся с полипиримидиновым трактом, стимулирует IRES полиовируса, модулируя связывание eIF4G.EMBO J. 2010; 29 (21): 3710–22.
CAS PubMed PubMed Central Статья Google ученый
Кафасла П., Лин Х, Карри С., Джексон Р.Дж. Активация пикорнавирусных IRES с помощью PTB показывает дифференциальную зависимость от каждого РНК-связывающего домена PTB. РНК. 2011. 17 (6): 1120–31.
CAS PubMed PubMed Central Статья Google ученый
Шон П., Нгуен Дж. Х., Семлер Б.Измененные взаимодействия между стеблем-петлей IV в 5′-некодирующей области РНК вируса Коксаки и поли (rC) связывающим белком 2: влияние на IRES-опосредованную трансляцию и вирусную инфекционность. Вирусология. 2009. 389 (1–2): 45–58.
CAS PubMed PubMed Central Статья Google ученый
Choi K, Kim JH, Li X, Paek KY, Ha SH, Ryu SH, et al. Идентификация клеточных белков, усиливающих активность внутренних участков входа в рибосомы, путем конкуренции с олигодезоксинуклеотидами.Nucleic Acids Res. 2004. 32 (4): 1308–17.
CAS PubMed PubMed Central Статья Google ученый
Луо З, Донг Х, Ли И, Чжан Кью, Ким С, Сонг Й и др. PolyC-связывающий белок 1 взаимодействует с 5′-нетранслируемой областью РНК энтеровируса 71 в мембранно-ассоциированном комплексе для облегчения репликации вируса. PLoS One. 2014; 9 (1): e87491.
PubMed PubMed Central Статья CAS Google ученый
Zhang H, Song L, Cong H, Tien P. Ядерный белок Sam68 взаимодействует с внутренним сайтом входа в рибосомы Enterovirus 71 и положительно регулирует трансляцию вирусного белка. J Virol. 2015. 89 (19): 10031–43.
CAS PubMed PubMed Central Статья Google ученый
Su YS, Tsai AH, Ho YF, Huang SY, Liu YC, Hwang LH. Стимуляция трансляции энтеровируса 71, зависящей от внутреннего сайта входа в рибосомы (IRES), с помощью DDX3X РНК-геликазы и вирусных протеаз 2A и 3C.Front Microbiol. 2018; 9: 1324.
PubMed PubMed Central Статья Google ученый
Лин Дж.Й., Брюэр Дж., Ли М.Л. HuR и Ago2 связывают внутренний сайт входа в рибосомы энтеровируса 71 и способствуют трансляции и репликации вируса. PLoS One. 2015; 10 (10): e0140291.
PubMed PubMed Central Статья CAS Google ученый
Huang PN, Lin JY, Locker N, Kung YA, Hung CT, Lin JY и др.Белок 1, связывающий далеко расположенный выше элемент, связывает внутренний сайт входа в рибосому энтеровируса 71 и усиливает трансляцию вируса и рост вируса. Nucleic Acids Res. 2011. 39 (22): 9633–48.
CAS PubMed PubMed Central Статья Google ученый
Лин Дж.Й., Ли М.Л., Ши СР. Белок 2, связывающий далеко расположенный выше элемент, взаимодействует с внутренним сайтом входа в рибосомы энтеровируса 71 и негативно регулирует вирусную трансляцию.Nucleic Acids Res. 2009. 37 (1): 47–59.
CAS PubMed Статья PubMed Central Google ученый
Каткарт А.Л., Розович Дж.М., Семлер Б.Л. Белок распада мРНК клеток AUF1 отрицательно регулирует инфекции энтеровирусов и риновирусов человека. J Virol. 2013. 87 (19): 10423–34.
CAS PubMed PubMed Central Статья Google ученый
Ульмер В., Семлер Б.Л.Прямые и косвенные эффекты на трансляцию вирусов и репликацию РНК необходимы для ограничения AUF1 энтеровирусных инфекций в клетках человека. MBio. 2018; 9 (5): e01669.
PubMed PubMed Central Статья Google ученый
Holcik M, Liebhaber SA. Четыре высокостабильных эукариотических мРНК собирают 3′-нетранслируемые области РНК-белковые комплексы, разделяющие цис- и транс-компоненты. Proc Natl Acad Sci. 1997. 94 (6): 2410–4.
CAS PubMed Статья PubMed Central Google ученый
Blyn LB, Chen R, Semler BL, Ehrenfeld E. Связывание белков клетки-хозяина с доменом IV 5′-некодирующей области РНК полиовируса. J Virol. 1995. 69 (7): 4381–9.
CAS PubMed PubMed Central Статья Google ученый
Blyn LB, Swiderek KM, Richards O, Stahl DC, Semler BL, Ehrenfeld E. Поли (rC) связывающий белок 2 связывается со стеблевой петлей IV 5′-некодирующей области 5′-некодирующей области РНК полиовируса: идентификация с помощью автоматизированной жидкости хроматография-тандемная масс-спектрометрия.Proc Natl Acad Sci U S. A. 1996; 93 (20): 11115–20.
CAS PubMed PubMed Central Статья Google ученый
Bedard KM, Daijogo S, Semler BL. Нуклео-цитоплазматический белок SR участвует в инициации вирусной IRES-опосредованной трансляции. EMBO J. 2007; 26 (2): 459–67.
CAS PubMed Статья PubMed Central Google ученый
Фицджеральд К.Д., Семлер Б.Л.Повторная локализация клеточного белка SRp20 во время полиовирусной инфекции: соединение вирусного IRES с аппаратом трансляции клетки-хозяина. PLoS Pathog. 2011; 7 (7): e1002127.
CAS PubMed PubMed Central Статья Google ученый
Дэн Х, Ван Кью, Йи Л, Лу Дж, Цзяо Й, Ли Х и др. Hsp27 отвечает и облегчает репликацию энтеровируса A71 за счет усиления вирусной IRES-опосредованной трансляции. J Virol. 2019; 93: 02322–18.
Артикул Google ученый
Dong Q, Men R, Dan X, Chen Y, Li H, Chen G, et al. Hsc70 регулирует активность IRES и служит противовирусной мишенью для инфекции энтеровируса A71. Antivir Res. 2018; 150: 39–46.
CAS PubMed Статья PubMed Central Google ученый
Леонг С.И., Онг Б.К., Чу Дж.Дж. Роль деформированной NCK-related kinase (MINK), новой киназы семейства Ste20, в IRES-опосредованной трансляции белков энтеровируса человека 71.PLoS Pathog. 2015; 11 (3): e1004686.
PubMed PubMed Central Статья CAS Google ученый
Jang SK, Wimmer E. Cap-независимая трансляция РНК вируса энцефаломиокардита: структурные элементы внутреннего сайта входа в рибосомы и участие клеточного РНК-связывающего белка 57 кДа. Genes Dev. 1990; 4 (9): 1560–72.
CAS PubMed Статья PubMed Central Google ученый
Blyn LB, Towner JS, Semler BL, Ehrenfeld E. Требование поли (rC) связывающего белка 2 для трансляции РНК полиовируса. J Virol. 1997. 71 (8): 6243–6.
CAS PubMed PubMed Central Статья Google ученый
Лефферс Х., Дейгаард К., Селис Дж. Характеристика двух основных клеточных поли (rC) -связывающих белков человека, каждый из которых содержит три K-гомологичных (KH) домена. Eur J Biochem. 1995. 230 (2): 447–53.
CAS PubMed Статья PubMed Central Google ученый
Sanford JR, Gray NK, Beckmann K, Cáceres JF. Новая роль челночных белков SR в трансляции мРНК. Genes Dev. 2004. 18 (7): 755–68.
CAS PubMed PubMed Central Статья Google ученый
Hung CT, Kung YA, Li ML, Brewer G, Lee KM, Liu ST и др. Аддитивное стимулирование трансляции, опосредованной внутренним сайтом входа в рибосомы вируса, с помощью белка 1, связывающего элементы, расположенного далеко выше по течению, и продукта расщепления, индуцированного энтеровирусом 71.PLoS Pathog. 2016; 12 (10): e1005959.
PubMed PubMed Central Статья CAS Google ученый
Санс М.А., Велновска Э., Редондо Н., Карраско Л. Трансляции, управляемой пикорнавирусом. IRES затрудняется репликонами вируса Синдбис: спасение с помощью протеазы полиовируса 2А. J Mol Biol. 2010. 402 (1): 101–17.
CAS PubMed Статья PubMed Central Google ученый
Stricher F, Macri C, Ruff M, Muller S. Белок-шаперон HSPA8 / HSC70: структура, функция и химическое нацеливание. Аутофагия. 2013; 9 (12): 1937–54.
CAS PubMed Статья PubMed Central Google ученый
Бёкинг Т., Агует Ф., Харрисон С.К., Кирххаузен Т. Одномолекулярный анализ молекулярной дизассемблазы раскрывает механизм удаления покрытия клатрина под действием Hsc70. Nat Struct Mol Biol. 2011; 18 (3): 295.
PubMed PubMed Central Статья CAS Google ученый
Chuang C-K, Ян Т-Х, Чен Т-Х, Ян Ц-Ф, Чен В-Дж. Изоформа D родственного белка теплового шока 70 необходима для клатрин-зависимого эндоцитоза вируса японского энцефалита в клетках C6 / 36. J Gen Virol. 2015; 96 (4): 793–803.
CAS PubMed Статья PubMed Central Google ученый
Lai MC, Chang WC, Shieh SY, Tarn WY. DDX3 регулирует рост клеток посредством контроля трансляции циклина E1. Mol Cell Biol. 2010. 30 (22): 5444–53.
CAS PubMed PubMed Central Статья Google ученый
Lai MC, Lee YH, Tarn WY. Геликаза DDX3 РНК DEAD-бокса связывается с рибонуклеопротеинами экспортного мессенджера, а также с концевым белком и участвует в контроле трансляции. Mol Biol Cell. 2008. 19 (9): 3847–58.
CAS PubMed PubMed Central Статья Google ученый
Хо Би Си, Ю С-Л, Чен Дж.Дж., Чанг С-И, Ян Би С, Хун Ц. С. и др. Индуцированная энтеровирусом miR-141 способствует отключению трансляции белка-хозяина, воздействуя на фактор инициации трансляции eIF4E. Клеточный микроб-хозяин. 2011; 9 (1): 58–69.
CAS PubMed Статья PubMed Central Google ученый
Weng K-F, Hung C-T, Hsieh P-T, Li M-L, Chen G-W, Kung Y-A, et al. Цитоплазматический РНК-вирус генерирует функциональные вирусные малые РНК и регулирует вирусную активность IRES в клетках млекопитающих.Nucleic Acids Res. 2014. 42 (20): 12789–805.
CAS PubMed PubMed Central Статья Google ученый