
Состав и функции крови — урок. Биология, Человек (8 класс).
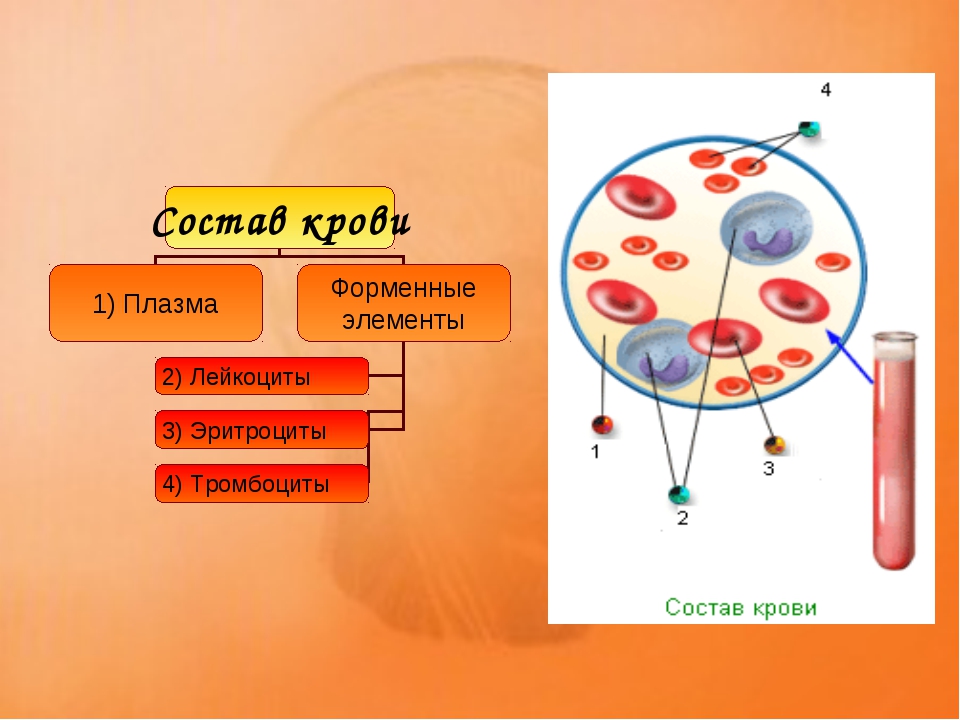
Общий объём крови у взрослого человека составляет \(4\)— \(6\) л. \(55\)–\(60\) % приходится на жидкую часть — плазму, а \(40\)–\(45\) % — на форменные элементы: эритроциты, лейкоциты и тромбоциты.
Рис. \(1\). Состав крови
Больше всего в плазме воды — до \(90\) %. В воде растворены минеральные соли и органические соединения. Часть этих веществ — питательные вещества, переносимые кровью к различным органам.
Состав плазмы не меняется, несмотря на постоянное поступление в кровь многих веществ. Выведение избытка веществ происходит через органы дыхания и выделения. Лёгкие забирают из крови избыток углекислого газа, а почки удаляют с мочой лишнюю воду, избыток минеральных солей и опасные продукты обмена.
Рис. \(2\). Состав плазмы
Образование форменных элементов крови происходит в красном костном мозге из стволовых клеток
 Красный костный мозг — это кроветворный орган, который находится между костными пластинками губчатого вещества костей. Его масса у взрослого человека составляет около \(4\) % от веса тела.
Красный костный мозг — это кроветворный орган, который находится между костными пластинками губчатого вещества костей. Его масса у взрослого человека составляет около \(4\) % от веса тела.
Рис. \(3\). Образование форменных элементов крови
К кроветворным органам относятся также лимфатические узлы, селезёнка и др.
Функции крови:
- дыхательная — переносит кислород от лёгких ко всем клеткам организма и углекислый газ — в обратном направлении.
- Питательная — переносит питательные вещества, которые всасываются в кишечнике.
- Выделительная
- Терморегуляционная — при пониженной температуре окружающей среды кровь, нагреваясь, переносит тепло из скелетных мышц и печени к тем органам, которые необходимо согреть (кожа, мозг и др.).
- Защитная — благодаря лимфоцитам и антителам уничтожаются и нейтрализуются попадающие внутрь организма опасные микробы и вещества; тромбоциты обеспечивают свёртываемость крови.
- Регуляторная — кровь транспортирует по организму гормоны и другие вещества и обеспечивает гомеостаз (постоянство внутренней среды организма).
Все перечисленные функции крови обусловлены её способностью переносить вещества от одних органов к другим и поэтому их можно объединить в одну функцию — транспортную.
Источники:
Рис. 1. Состав крови: https://image.shutterstock.com/image-vector/vector-diagram-blood-composition-educational-600w-1687292392.jpg
Рис. 2. Состав плазмы: © ЯКласс
Рис. 3. Образование форменных элементов крови: https://image.shutterstock.com/image-vector/blood-stem-cells-haematopoiesis-editable-600w-1233232918.jpg
Процедура, алгоритм взятия крови из вены и другого биологического материала
Безопасность пациентов – приоритет нашей компании. Для взятия биологического материала мы используем только одноразовые стерильные системы и инструменты, соответствующие мировым стандартам качества.
Вакуумные системы взятия крови Vacuette
Использование вакуумных систем взятия крови повышенной безопасности Vacuette, Greiner Bio-One, Австрия, полностью исключает контакт с кровью, обеспечивая безопасность как пациента, так и персонала. Кроме того, система обеспечивает сохранность крови при транспортировке, получение качественного материала для исследования (сыворотка, плазма), минимизирует время подготовки проб к исследованию. В нашей лаборатории во всех автоматических анализаторах именно первичная пробирка, в которую брали кровь пациенту, используется для производства анализа.
Вакуумные системы Vacuette соответствуют требованиям и рекомендациям международных стандартов ISO 6710 (EN 14820) «Одноразовые контейнеры для взятия проб венозной крови» и Института Клинических и Лабораторных Стандартов (LLSI).
Механизм забора крови вакуумной системой заключается в том, что под действием вакуума кровь поступает через иглу напрямую из вены в пробирку, обеспечивая четкое дозирование крови и возможность набрать кровь в 2 и более пробирки за очень короткий промежуток времени без повторного введения иглы в вену. Процедура проводится медицинскими сестрами, имеющими многолетний опыт работы с венами любой сложности, занимает несколько секунд, безболезненна, абсолютно безопасна. Вскрытие системы проводится в присутствии клиента, повторное использование вакуумных систем взятия крови невозможно. Система штрих-кодирования пробирок обеспечивает четкую идентификацию проб и исключает подмену образцов.
Процедура проводится медицинскими сестрами, имеющими многолетний опыт работы с венами любой сложности, занимает несколько секунд, безболезненна, абсолютно безопасна. Вскрытие системы проводится в присутствии клиента, повторное использование вакуумных систем взятия крови невозможно. Система штрих-кодирования пробирок обеспечивает четкую идентификацию проб и исключает подмену образцов.
Почему лучше сдавать кровь на общий анализ из вены, а не из пальца?
Венозная кровь – лучший материал не только для определения биохимических, гормональных, иммунологических и серологических показателей, но и для общеклинического исследования крови. Современные анализаторы, с помощью которых проводят подсчет клеток крови, определение гемоглобина, гематокрита, СОЭ и других показателей, стандартизированы и сертифицированы для работы с венозной кровью. Выпускаемые фирмами-производителями калибровочные и контрольные материалы предназначены для подготовки к работе и контроля за работой анализаторов по венозной крови.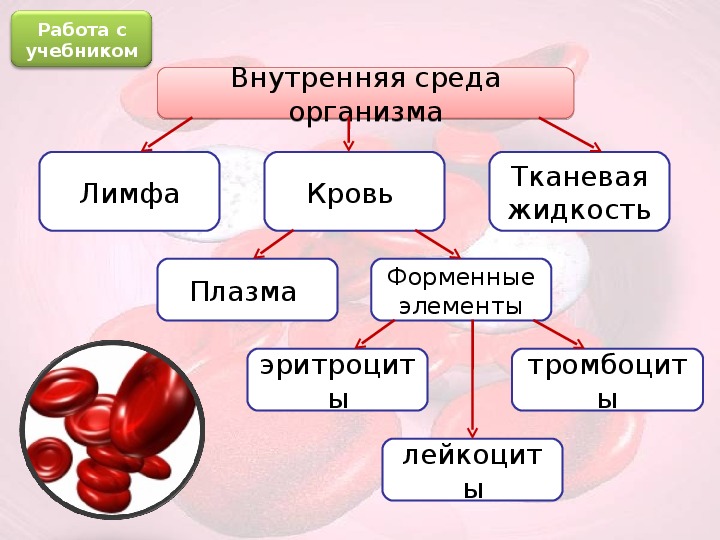
При взятии крови из пальца крайне сложно соблюсти стандартизацию процедуры, а необходимость разведения образцов приводит к значительным разбросам получаемых результатов, и, как следствие, необходимости повторного взятия и исследования для уточнения результата. Для общеклинического исследования кровь из пальца рекомендуется брать в следующих случаях:
- при ожогах, занимающих большую площадь поверхности тела пациента;
- при выраженном ожирении;
- при установленной склонности к венозным тромбозам.
Взятие биологического материала для бактериологических исследований (отделяемое дыхательных путей, мочеполового тракта)
Проводится с использованием современных систем премиум-класса с транспортной средой Эймса, обеспечивающих сохранность микроорганизмов и качественное проведение всех этапов анализа, связанных с диагностикой инфекционных процессов от определения микроорганизма, вызывающего заболевание, до подбора эффективного препарата для лечения.
Правила сложения: что можно узнать о будущем ребенке по своей группе крови | Статьи
Руководитель департамента по развитию медицины BestDoctor Юлия Ткаченко сообщила 30 августа «Известиям», что первая группа крови является универсальным донором, то есть кровь этой группы можно переливать любым пациентам. Закон, однако, предписывает переливать кровь только пациентам с идентичными группами. Что такое группа крови и какие подробности о будущем ребенке можно узнать по группам крови родителей, разбирались «Известия».
Сколько бывает групп крови
Группа крови — один из важнейших факторов, определяющих биологические данные нашего организма. В частности, знать свою группу крови следует на случай необходимости переливания или срочного донорства. Это не меняющийся в течение жизни признак, передаваемый по наследству. Всего существуют четыре группы крови, определяемых зависимости от наличия на поверхности эритроцитов тех или иных белков, называемых аглютиногенами, и белков, находящихся в плазме, — аглютининов.
Фото: РИА Новости/Владимир Песня
«В настоящее время по закону переливать кровь можно только с идентичными группами (первую — первой, вторую — второй и т.д.). Но если исходить из биологии, то первая группа крови является универсальным донором (ее можно переливать любой другой группе), а четвертая — универсальным реципиентом (человеку с четвертой группой можно перелить любую из групп)», — объяснила «Известиям» руководитель департамента по развитию медицины BestDoctor Юлия Ткаченко.
К открытию групп крови привели в конце XIX века исследования австрийца Карла Ландштайнера. Изучая эритроциты — красные кровяные клетки, ученый обнаружил определенные закономерности. Так, в образцах крови одних пациентов Ландштайнер выделил особый маркер (он обозначил его буквой А,чем и положил начало будущей классификации).
Как наследуется группа крови
Как уже говорилось, группа крови — наследуемый признак и передается с генами родителей потомству. Таким образом, ребенок, по идее, должен наследовать группу крови одного из родителей. Однако не всё так просто. Поскольку открытые еще Грегором Менелем рецессивные и доминантные признаки при наследовании распространяются и на кровь, ребенок имеет несколько вариантов группы крови. Наибольшее число таких вариантов у детей родителей с группами крови 0 и АВ.
При этом, по данным науки, в семье, где у одного из родителей группа крови АВ, никогда не может родиться ребенок с группой крови 0, какая бы группа ни была у второго родителя. Это правило работает и в обратную сторону: если у отца или матери первая группа, то рождение детей с кровью АВ в таком браке невозможно. С остальными группами крови вариации можно для простоты свести в таблицу:
Автор цитаты
• У родителей с первой группой крови может родиться ребенок только с первой группой.
• У родителей со второй — ребенок с первой или второй.
• У родителей с третьей — ребенок с первой или третьей.
• У родителей с первой и второй — ребенок с первой или второй.
• У родителей с первой и третьей — ребенок с первой или третьей.
• У родителей с второй и третьей — ребенок с любой группой крови.
• У родителей с первой и четвертой — ребенок с второй и третьей.
• У родителей с второй и четвертой — ребенок с второй, третьей и четвертой
• У родителей с третьей и четвертой — ребенок с второй, третьей и четвертой.
• У родителей с четвертой — ребенок с второй, третьей и четвертой.

При определении группы крови у новорожденного врачи учитывают, что антиген В полностью проявляется лишь к концу первого года жизни. Таким образом, ребенок с третьей группой крови может при рождении получить в медицинские документы первую, а с четвертой — вторую. По мере взросления ребенка это недоразумение обычно устраняется.
Зачем нужен резус-фактор
Кроме группы, кровь определяется и таким важным признаком, как резус-фактор (Rh). Он может быть положительным или отрицательным и обозначается, соответственно, знаками (+) и (-). Резус-фактор зависит от наличия или отсутствия антигена D на поверхности эритроцитов. Назначение этого антигена до сих пор до конца не установлено, однако известно, что 85% населения Земли — резус-положительны. Несовпадение резус-фактора при беременности или переливании крови может привести к тяжким последствиям, так как соприкосновение крови с разными значениями Rh начинают вырабатываться антитела.
Фото: РИА Новости/Георгий Зимарев
Однако опасна лишь ситуация, при которой резус-отрицательная мать вынашивает резус-положительного ребенка. При обратном соотношении резус-конфликта не происходит. Наибольшая опасность возникает в ситуации, когда резус-отрицательными являются оба родителя. В любом случае паре, задумывающейся о продолжении рода, всегда необходимо обратиться к медикам для консультации.
Урок 15. состав крови. постоянство внутренней среды – Биология – 8 класс
Конспект
Кровь – жидкая соединительная ткань. Большую ее часть составляет межклеточное вещество – плазма крови. На ее долю приходится 55% от объема крови.
Основную часть плазмы составляет вода. Ее там до 92%. Также в состав плазмы обязательно входят белки – 7-8% (например, фибриноген), минеральные соли (хлорид натрия) – 0,9%, жироподобные вещества – 0,7%, глюкоза – 0,12 %, витамины и азотистые вещества.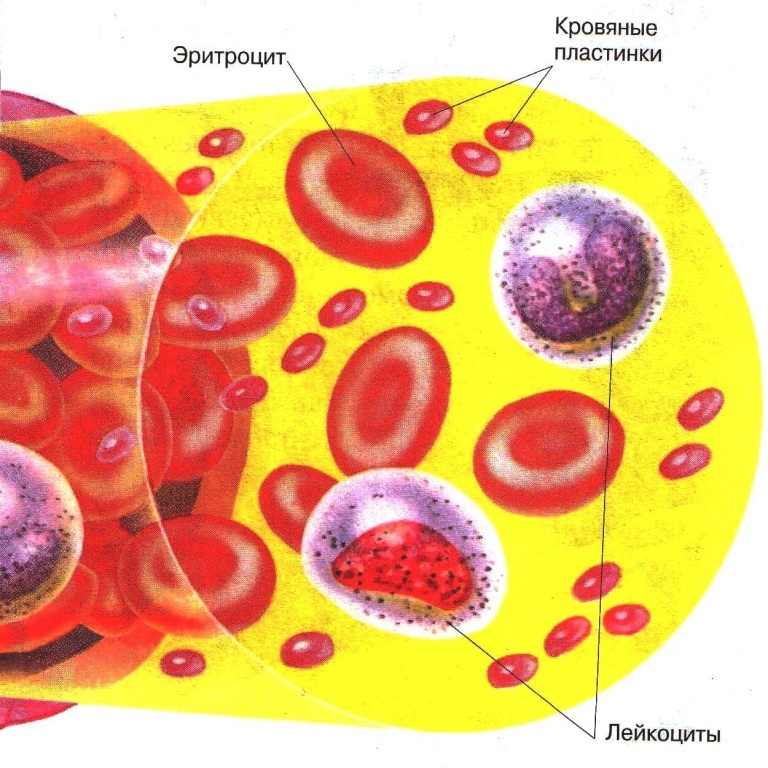 Постоянное содержание минеральных солей и глюкозы в плазме крови является обязательным условием нормальной жизнедеятельности клеток организма.
Постоянное содержание минеральных солей и глюкозы в плазме крови является обязательным условием нормальной жизнедеятельности клеток организма.
Форменные элементы крови – вторая составная часть кровь. Среди них выделяют тромбоциты, эритроциты и лейкоциты.
Кровь человека красная, потому что в ней больше всего именно красных кровяных телец – эритроцитов. В 1 кубическом миллиметре крови их до 5 миллионов. Их функция – транспорт молекул кислорода из легких к клеткам организма и частично молекул углекислого газа в обратном направлении. Выполнение этой функции возможно благодаря наличию в эритроцитах красного железосодержащего белка – гемоглобина. Зрелые эритроциты млекопитающих животных и человека имеют форму двояковогнутого диска и не содержат ядра (ядро присутствует у молодых эритроцитов). Такие особенности строения позволяют разместить внутри эритроцита больше молекул гемоглобина и более эффективно осуществлять газообмен. Срок жизни эритроцитов – 3-4 месяца. Старые эритроциты разрушаются в печени или селезенке, новые эритроциты образуются в красном костном мозге.
Лейкоциты – единственные из форменных элементов крови, которые с полным правом могут называться полноценными клетками. В кубическом миллиметре крови их примерно в тысячу раз меньше, чем эритроцитов. Форма непостоянная, могут передвигаться благодаря ложноножкам, в том числе и вне сосудов. Основная функция – защитная – обеспечение иммунитета, осуществляется по-разному. Есть лейкоциты, поглощающие и переваривающие инородные тела путем фагоцитоза. Данное явление было открыто русским ученым Ильей Ильичем Мечниковым. Другие лейкоциты выделяют особые защитные белки – антитела, нейтрализующие чужеродные белки – антигены.
Тромбоциты, или кровяные пластинки также, как и другие форменные элементы, образуются в красном костном мозге. Их содержание в кубическом миллиметре от 200 до 400 тысяч. Обеспечение свертывания крови – их главная функция.
Общий объем крови в организме мужчины примерно 5 л, в организме женщины – 4,5 л. Часть крови находится в так называемых депо крови – сосудах кожи, печени и селезенки. В случае кровопотери или большой физической нагрузки кровь из депо поступает в кровоток.
В случае кровопотери или большой физической нагрузки кровь из депо поступает в кровоток.
Относительно постоянный химический состав плазмы крови – один из самых ярких примеров гомеостаза. Показателен пример с хлоридом натрия – в плазме крови его содержание 0,9%. В случае уменьшения этой величины вода из плазмы начинает поступать внутрь эритроцитов, и они лопаются; в случае увеличения – эритроциты теряют воду, сморщиваются и погибают, образую при этом тромбы.
Такая же ситуация с необходимостью поддержания относительного постоянства уровня глюкозы. При падении содержания глюкозы до 0,05% возможна потеря сознания и гибель. Клеткам головного мозга в этом случае не хватает источника энергии для работы.
Подведем итог.
Кровь – жидкая соединительная ткань, на 55% состоящая из межклеточного вещества – плазмы крови с большим количеством важных неорганических и органических веществ и на 45% – из форменных элементов крови – лейкоцитов, тромбоцитов и эритроцитов.
Всё о сердце | Ассоциация сердечно-сосудистых хирургов России Секция “Кардиология и визуализация в кардиохирургии”
СЕРДЦЕ, КАК ОНО РАБОТАЕТ?
Сердце человека – это сильный мышечный насос. Каждый день сердце сокращается и расслабляется 100000 раз и перекачивает 7600 литров крови. За 70 лет жизни среднестатистическое человеческое сердце сокращается более 2,5 миллионов раз.
Каждый день сердце сокращается и расслабляется 100000 раз и перекачивает 7600 литров крови. За 70 лет жизни среднестатистическое человеческое сердце сокращается более 2,5 миллионов раз.
Сердце прокачивает кровь через кровеносную систему. Кровеносная система – это сеть эластических трубок, по которым кровь поступает к органам и тканям организма. В состав кровеносной системы входят сердце и сосуды: артерии, артериолы, капилляры (самые мелкие сосуды), венулы и вены. Артерии несут богатую кислородом кровь ко всем частям тела. Вены несут углекислый газ и продукты распада обратно к сердцу и легким. Если все сосуды человеческого тела соединить вместе и вытянуть в одну линю, они покроют расстояние в 96,5 тысяч километров. Этого будет достаточно, чтобы обхватить землю более чем 2 раза. Кровь несет кислород и питательные вещества ко всем органам и тканям, включая само сердце. Из тканей в кровь поступают продукты обмена. Продукты обмена удаляются почками, печенью и легкими.
Сердце состоит из 4 камер; 2 предсердия и 2 желудочка. Камеры разделены клапанами, которые открываются и закрываются в процессе сокращения сердца, позволяя крови течь только в одном направлении. Клапаны открываются, когда при сокращении сердца давление в камерах повышается.
Камеры разделены клапанами, которые открываются и закрываются в процессе сокращения сердца, позволяя крови течь только в одном направлении. Клапаны открываются, когда при сокращении сердца давление в камерах повышается.
Клапаны сердца:
– Трикуспидальный клапан между правым предсердием и правым желудочком
– Легочный клапан между правым желудочком и легочной артерией
– Митральный клапан между левым предсердием и левым желудочком
– Аортальный клапан между левым желудочком и аортой
Каждый клапан имеет несколько створок. У митрального клапана 2 створки, у других клапанов 3.
Как работает сердце?
Сердце качает кровь за счет согласованного последовательного сокращения его камер. В правое предсердие кровь попадает из вен. Венозная кровь богата углекислым газом и почти не содержит кислорода. По сравнению с артериальной кровью она имеет более темный цвет. Когда сердечная мышца расслабляется, венозная кровь поступает через открытый трикуспидальный клапан в правый желудочек.
Электрический импульс, запускает сердечное сокращение, которое начинается с сокращения предсердий. Правое предсердие, сокращаясь, заполняет правый желудочек дополнительным объемом крови. После сокращений правого предсердия сокращается правый желудочек. В этот момент трикуспидальный клапан закрывается, не давая крови поступать обратно в предсердие, и вся кровь из правого желудочка поступает в легочную артерию и затем в легкие. В легких из крови освобождается углекислый газ и кровь насыщается кислородом. Богатая кислородом артериальная кровь поступает из легких в левое предсердие.
Синхронно с правым предсердием сокращается левое предсердие. Из него кровь через митральный клапан поступает в левый желудочек. Левый желудочек, сокращаясь, толкает кровь через аортальный клапан в аорту. От аорты отходят множество артерий, несущих кровь ко всем органам и тканям.
Все четыре камеры сердца должны сокращаться определенным образом. Сокращениями сердца управляют электрические импульсы. Камеры сердца начинают сокращаться, после того как через них проходят электрические импульсы. Импульсы зарождаются в особой части нервной системы сердца, называемой синусовым узлом. Синусовый узел является основным водителем ритма, заставляющим сердце сокращаться. Водитель ритма регенерирует импульсы с определенной частотой. Эмоциональные реакции и гормональные воздействия могут изменять это частоту, заставляя сердце биться чаще или реже.
Камеры сердца начинают сокращаться, после того как через них проходят электрические импульсы. Импульсы зарождаются в особой части нервной системы сердца, называемой синусовым узлом. Синусовый узел является основным водителем ритма, заставляющим сердце сокращаться. Водитель ритма регенерирует импульсы с определенной частотой. Эмоциональные реакции и гормональные воздействия могут изменять это частоту, заставляя сердце биться чаще или реже.
Электрический импульс, возникающий в синусовом узле, идет через правое и левое предсердие, заставляя мышечные клетки сокращаться. После того, как предсердия сократились, электрический импульс проходит по нервной системе сердца дальше к желудочкам, заставляя их сокращаться и изгонять кровь в сосуды. Роль электрического импульса заключается в обеспечении координированного сокращения сердца, необходимого для его хорошей работы.
Материал подготовлен Голуховой Е.З.
Пути передачи ВИЧ-инфекции — Как обезопасить себя? — ВИЧ/СПИД: материалы Детского фонда ООН
Как обезопасить себя?
Пути передачи ВИЧ-инфекции
ВИЧ передается при передаче биологических жидкостей, содержащих вирус иммунодефицита, от одного человека другому.

Вирус иммунодефицита присутствует в следующих биологических жидкостях: кровь, сперма и пре-эякуляционные выделения, вагинальные выделения и грудное молоко. Данный вирус также содержится в слюне, однако, по мнению экспертов, его концентрация слишком мала для того, чтобы вызвать заболевание.
Встреча в джаз-клубе в столице Замбии Лусаке,. Фото ЮНИСЕФ.Отсюда следует, что поцелуи, даже глубокие поцелуи с открытым ртом, не несут никакого риска. Также невозможно заразиться ВИЧ при обычных прикосновениях, например при рукопожатии. Более того, нельзя заразиться ВИЧ от прикосновения к той вещи, которой уже касался ВИЧ-инфицированный. ВИЧ также невозможно заразиться воздушным путем. Биологические жидкости, содержащие вирус иммунодефицита, попадают в организм человека только через повреждения в тканях/коже.
Секс
Незащищенный проникающий секс является наиболее распространенным способом попадания биологических жидкостей из одного организма в другой, что может повлечь распространение вируса. «Незащищенный» означает секс без презерватива, «проникающий» значит, что во время полового акта пенис проникает или входит во влагалище или анальное отверстие. В этом случае сперма, зараженная вирусом иммунодефицита, вагинальная жидкость или кровь одного человека могут попасть в организм другого человека через мельчайшие порезы или раны во влагалище, анальном отверстии или на пенисе. Данные порезы или раны могут быть настолько малы, что их невозможно увидеть.
«Незащищенный» означает секс без презерватива, «проникающий» значит, что во время полового акта пенис проникает или входит во влагалище или анальное отверстие. В этом случае сперма, зараженная вирусом иммунодефицита, вагинальная жидкость или кровь одного человека могут попасть в организм другого человека через мельчайшие порезы или раны во влагалище, анальном отверстии или на пенисе. Данные порезы или раны могут быть настолько малы, что их невозможно увидеть.
Наличие другого венерического заболевания повышает вероятность заражения ВИЧ-инфекцией, т.к. венерические заболевания вызывают повреждения тканей половых органов, способствуя попаданию зараженной крови в кровеносную систему. Венерические заболевания могут также становиться причиной серьезных проблем со здоровьем. Поэтому при возникновении в половых органах таких симптомов, как боль, язвы или раны, зуд или опухание, проблемы с мочеиспусканием, следует обязательно обратиться к медицинскому работнику. Все больницы, медицинские центры, клиники и частные врачи могут вылечить венерические заболевания. Не следует стесняться: вне зависимости от того, где проходит лечение, вся информация об этом является конфиденциальной.
Не следует стесняться: вне зависимости от того, где проходит лечение, вся информация об этом является конфиденциальной.
Наркотики и иглы
Наркоманы, использующие нестерильные иглы, шприцы и воду для промывки, подвергают себя огромному риску заражения ВИЧ-инфекцией. Биологические жидкости (чаще всего кровь), содержащие вирус иммунодефицита, могут попасть прямо в систему кровообращения через иголки на шприце, что делает риск заражения очень высоким.
Татуировки и пирсинг, производимые с использованием нестерильного оборудования, также несут риск заражения ВИЧ-инфекцией.
Передача инфекции от матери ребенку
Матери, зараженные ВИЧ-инфекцией, могут передать ее своим детям во время беременности, родов или в период грудного вскармливания. Если ВИЧ-инфицированная женщина становится беременной, существует 35-процентная вероятность того, что она передаст вирус ребенку в случае отсутствия превентивного лечения.
Переливание крови и препараты крови
Каждому донору крови следует пройти тест на ВИЧ, чтобы зараженная кровь не попала в другой организм. К сожалению, не везде медицинские услуги включают проверку донорской крови на содержание вируса иммунодефицита.
К сожалению, не везде медицинские услуги включают проверку донорской крови на содержание вируса иммунодефицита.
Что такое тромбоз – венозный и артериальный тромбозы
За жизнедеятельность нашего организма отвечают множество биологических систем. Одна из них – гемостаз. Его задача – сохранять кровь в жидком состоянии, останавливать кровотечения в случае повреждения стенок сосудов, а также растворять тромбы. Тромбообразование – один из механизмов гемостаза. Образование тромба – это защитная реакция организма, когда нужно остановить кровотечение. При повреждении образуется кровяной сгусток (тромб), который, как заплатка, закрывает поврежденный участок, т.е. защищает организм от кровопотери. На языке медиков этот процесс называется тромбоз. [1]
С одной стороны, тромбоз защищает организм от больших кровопотерь. С другой, может спровоцировать
тяжелые заболевания, связанные с нарушением кровотока. Это инсульт, инфаркт миокарда, ишемические повреждения органов
брюшной полости и забрюшинного пространства, гангрены конечностей. [2]
[2]
До середины 19 века тромбоз был загадкой для врачей, они пытались понять, что же это такое. Разгадать тайну удалось немецкому патологоанатому Рудольфу Вирхову. Он изучил механизмы возникновения тромбоза, выявив три базовых принципа. С тех пор прошло полтора столетия, но открытие берлинского врача по-прежнему актуально. Эту теорию принято называть триадой Вирхова. [3]
Рудольф Вирхов сформулировал три основных причины возникновения тромбоза. Первая – ухудшение состояния стенок сосудов. Это могут быть последствия травмы или хирургического вмешательства. Вирхов заметил, что тромбы чаще всего образуются там, где повреждена внутренняя структура сосудов. Вторая причина – замедление кровотока. Тромбы скапливаются там, где меняется скорость потока крови. Изменение вязкости крови также провоцирует тромбоз. В «густой» крови тромбы образуются чаще, чем в «жидкой». [3]
Тромбоз бывает двух форм – венозный и артериальный. Названия подсказывают, где именно
образуются тромбы: в первом случае в венах, во втором – в артериях.
Венозный тромбоз относят к острым заболеваниям, причин для его возникновения много, основные – это нарушение структуры венозной стенки во время операции, травмы, лучевой и химиотерапии, замедление скорости кровотока, повышенная свертываемость крови. [4]
Артериальный тромбоз (атеротромбоз) – патологическое состояние, почти у всех больных есть атеросклеротические бляшки. Это холестериновые отложения, они суживают просвет в артерии и приводят к дефициту кровообращения. Со временем бляшки обрастают тромбом. [2]
Тромбоэмболия Легочной Артерии (ТЭЛА) – тяжелое, смертельно опасное заболевание, при котором происходит полное или частичное закрытие просвета легочной артерии тромбом. Как правило, ТЭЛА является осложнением тромбоза глубоких вен ног. [4]
Вы можете ознакомиться со списком и картой лечебно-профилактических учреждений, где возможно проконсультироваться с профильными специалистами.
Источники:
- Струкова С.
 М. Основы физиологии гемостаза (2-ое изд. переработанное и дополненное) – Москва: Изд.МГУ, 2013.- 186
с.
М. Основы физиологии гемостаза (2-ое изд. переработанное и дополненное) – Москва: Изд.МГУ, 2013.- 186
с. - Шиффман Ф. Дж. Патофизиология крови. Пер. с англ.- М.-СПб.: “Издательство БИНОМ” ¬ “Невский Диалект”, 2000.-448 с., ил.
- Нефедов В.П. и соавт. Практическая медицина 2011; 1(49):58-61
- Российские клинические рекомендации по диагностике, лечению и профилактике венозных тромбоэмболических осложнений (ВТЭО). Флебология, 2015: 4(2): 3-52
SARU.ENO.19.02.0227a
Нажмите на цифры для просмотра видео
4 Тромбоэмболия Легочной Артерии
Тромбоэмболия Легочной Артерии (ТЭЛА) – тяжелое, смертельно опасное заболевание, при котором происходит полное или частичное закрытие просвета легочной артерии тромбом. Как правило, ТЭЛА является осложнением тромбоза глубоких вен ног.
Назад1Сердечно-сосудистая система
Cердечно-сосудистая система – одна из важнейших систем организма,
обеспечивающих его жизнедеятельность. Сердечно-сосудистая система обеспечивает циркуляцию крови в организме
человека. Кровь с кислородом, гормонами и веществами по осудам разносится по организму.По пути она делится
этим богатством со всеми органами и тканями. Затем забирает все, что осталось от обмена веществ для дальнейшей
утилизации.
Сердечно-сосудистая система обеспечивает циркуляцию крови в организме
человека. Кровь с кислородом, гормонами и веществами по осудам разносится по организму.По пути она делится
этим богатством со всеми органами и тканями. Затем забирает все, что осталось от обмена веществ для дальнейшей
утилизации.
3 Тромбоз глубоких вен
Тромбоз глубоких вен возникает, когда тромб образуется в глубоких венах нижних конечностей.
Назад2 Тромбоз поверхностных вен
Тромбоз поверхностных вен обычно поражает поверхностные вены на ногах, иногда в паху.
НазадОбзор Крови
1. Кровь состоит на 55 % из плазмы и на 45 % из форменных элементов — эритроцитов, лейкоцитов и тромбоцитов.
Кровь представляет собой жидкую соединительную ткань, циркулирующую по всему телу. Почему считается, что это жидкая соединительная ткань, а не просто жидкость? Он состоит из живых клеток, взвешенных в плазме , жидкости, которая составляет около 55% крови. Плазма транспортирует клетки крови, белки, электролиты, гормоны и питательные вещества по всему телу.Он также приносит продукты жизнедеятельности из тканей организма в мочевыделительную систему, где почки фильтруют их из крови.
Почему считается, что это жидкая соединительная ткань, а не просто жидкость? Он состоит из живых клеток, взвешенных в плазме , жидкости, которая составляет около 55% крови. Плазма транспортирует клетки крови, белки, электролиты, гормоны и питательные вещества по всему телу.Он также приносит продукты жизнедеятельности из тканей организма в мочевыделительную систему, где почки фильтруют их из крови.
Существуют три широкие категории клеток крови, выполняющих важные функции. Эритроциты, или эритроцитов , переносят кислород из легких в остальные части тела. Лейкоциты, или лейкоцитов , помогают защитить организм от патогенов. Существует пять различных типов лейкоцитов, которые по-разному борются с инфекцией. Тромбоциты, или тромбоцитов , слипаются и образуют сгустки для восстановления разорванных кровеносных сосудов.
2. Кровь выполняет пять основных функций.
Кровь выполняет пять основных функций, которые делают ее необходимой для выживания человека:
- Кровь переносит кислород от легких к клеткам организма и доставляет углекислый газ от клеток организма к легким.
 Кислород является важным компонентом аэробного клеточного дыхания, осуществляемого клетками человеческого тела. Мы получаем кислород из вдыхаемого воздуха, а эритроциты переносят этот кислород из легких в остальные части тела.Углекислый газ образуется как побочный продукт клеточного дыхания и оседает в крови, поэтому его можно доставить в легкие и выдыхать.
Кислород является важным компонентом аэробного клеточного дыхания, осуществляемого клетками человеческого тела. Мы получаем кислород из вдыхаемого воздуха, а эритроциты переносят этот кислород из легких в остальные части тела.Углекислый газ образуется как побочный продукт клеточного дыхания и оседает в крови, поэтому его можно доставить в легкие и выдыхать. - Кровь переносит гормоны и питательные вещества по всему телу. Эндокринные железы, такие как гипофиз и надпочечники, выделяют гормоны в кровоток, который доставляет их к органам тела. Гормоны — это химические мессенджеры, которые регулируют многие функции организма. Когда мы перевариваем пищу, ворсинки тонкого кишечника всасывают питательные вещества в кровь.Эти важные молекулы — глюкоза, аминокислоты, витамины, минералы и жирные кислоты — помогают клеткам организма выживать и выполнять свои функции. Кровь также транспортирует отходы к почкам и печени, которые удаляют их и перерабатывают для выведения.

- Кровь регулирует температуру тела. Когда организму необходимо согреться или остыть, важную роль играет система кровообращения. Кровеносные сосуды в коже могут расширяться или сужаться, чтобы контролировать, сколько крови поступает на поверхность кожи.Расширение кровеносных сосудов приближает кровь к поверхности кожи, поэтому может выделяться тепло, помогающее охладить тело. Это называется расширением сосудов . Вазоконстрикция — это когда кровеносные сосуды сокращаются, удерживая кровь на большем расстоянии от поверхности кожи, чтобы предотвратить потерю тепла.
- Кровь защищает организм от патогенов. Лейкоциты играют ключевую роль во врожденных и адаптивных иммунных реакциях организма. Некоторые лейкоциты специализируются на поглощении бактерий и других патогенов посредством процесса, называемого фагоцитозом .Другие адаптировались для обнаружения и «маркировки» определенных патогенов для удаления.

- Сгустки крови для предотвращения кровопотери в местах повреждений. Когда кровеносный сосуд разрывается, тромбоциты в этой области активируются, соединяясь с другими тромбоцитами, образуя пробку для предотвращения дальнейшей потери крови. Эти тромбоциты выделяют ферменты, которые способствуют образованию тромбов.
3. Клетки крови образуются в красном костном мозге.
Красный костный мозг содержит гемопоэтические стволовые клетки, или гемоцитобласты , которые делятся и дифференцируются в миелоидные и лимфоидные стволовые клетки .
Миелоидные стволовые клетки дают начало эритроцитам, тромбоцитам и миелобластам — клеткам, которые дифференцируются в миелоидные лейкоциты: нейтрофилы, базофилы, эозинофилы и моноциты.
Лимфоидные стволовые клетки дают начало лимфобластам , которые дифференцируются в лейкоциты, классифицируемые как лимфоциты: В-клетки, Т-клетки и естественные клетки-киллеры (NK).
4. Существует пять типов лейкоцитов: нейтрофилы, базофилы, эозинофилы, моноциты и лимфоциты.
Биология и лечение плазмобластной лимфомы | Кровь
Исход у нелеченных пациентов с PBL неутешителен: медиана общей выживаемости составляет 3 месяца для ВИЧ-позитивных пациентов и 4 месяца для ВИЧ-отрицательных пациентов. 42 И наоборот, ВИЧ-положительные пациенты с ЛБЛ, достигшие полной ремиссии после химиотерапии, достигают лучших результатов. 3,44 Спонтанная регрессия PBL после начала кАРТ была отмечена в двух отчетах о случаях ВИЧ-позитивных пациентов. 45,46 Аналогично, у ВИЧ-отрицательного пациента с ревматоидным артритом была отмечена регрессия PBL после снижения дозы метотрексата. 47
Учитывая неблагоприятный прогноз для пациентов с PBL, мы можем утверждать, что не существует стандарта лечения пациентов с PBL. В частности, использование циклофосфамида, доксорубицина, винкристина и преднизолона (CHOP) считается неадекватной терапией, и современные руководства рекомендуют более интенсивные режимы. 48 Такие схемы включают инфузионное введение этопозида, винкристина и доксорубицина с болюсным введением циклофосфамида и преднизолона (EPOCH), 49 циклофосфамида, винкристина, доксорубицина, метотрексата, чередующегося с ифосфамидом, этопозидом и цитарабином (CODOX-M6). или гиперфракционированные циклофосфан, винкристин, доксорубицин и дексаметазон, чередующиеся с метотрексатом и цитарабином (гипер-CVAD). 51 Однако два исследования пациентов с PBL, получавших более интенсивные схемы химиотерапии, чем CHOP, не выявили улучшения выживаемости. 3,44 Метаанализ на уровне пациентов выявил преимущество в выживаемости при использовании EPOCH по сравнению с CHOP у пациентов с ВИЧ-ассоциированными лимфомами. 52,53 Однако неясно, были ли включены в этот анализ какие-либо пациенты с ЛПК.
В частности, использование циклофосфамида, доксорубицина, винкристина и преднизолона (CHOP) считается неадекватной терапией, и современные руководства рекомендуют более интенсивные режимы. 48 Такие схемы включают инфузионное введение этопозида, винкристина и доксорубицина с болюсным введением циклофосфамида и преднизолона (EPOCH), 49 циклофосфамида, винкристина, доксорубицина, метотрексата, чередующегося с ифосфамидом, этопозидом и цитарабином (CODOX-M6). или гиперфракционированные циклофосфан, винкристин, доксорубицин и дексаметазон, чередующиеся с метотрексатом и цитарабином (гипер-CVAD). 51 Однако два исследования пациентов с PBL, получавших более интенсивные схемы химиотерапии, чем CHOP, не выявили улучшения выживаемости. 3,44 Метаанализ на уровне пациентов выявил преимущество в выживаемости при использовании EPOCH по сравнению с CHOP у пациентов с ВИЧ-ассоциированными лимфомами. 52,53 Однако неясно, были ли включены в этот анализ какие-либо пациенты с ЛПК. Важно отметить, что два проспективных исследования оценивают безопасность и эффективность инфузионного EPOCH у пациентов с ДВККЛ высокого риска, включая лейкоциты (NCT010
Важно отметить, что два проспективных исследования оценивают безопасность и эффективность инфузионного EPOCH у пациентов с ДВККЛ высокого риска, включая лейкоциты (NCT010
Сообщалось лишь о нескольких случаях использования интратекальных агентов для сведения к минимуму риска поражения центральной нервной системы.Однако, учитывая высокую скорость пролиферации PBL, сильную связь с ВИЧ-инфекцией, высокую частоту экстранодального вовлечения и наличие транслокаций MYC, мы считаем, что интратекальную профилактику следует рассматривать у большинства пациентов с PBL. Улучшение результатов кАРТ среди пациентов с ВИЧ-ассоциированной ДВККЛ не было очевидным среди пациентов с ВИЧ-ассоциированной ЛБЛ. Два исследования показали, что ВИЧ-позитивные пациенты с PBL, которые также получали кАРТ, имели улучшенную выживаемость, 41,44 , тогда как другое исследование этого не сделало. 3 Информация о лучевой терапии довольно скудна. Лучевая терапия использовалась как часть лечения примерно в 20–30 случаях ЛБЛ, но на основании этого ограниченного опыта нельзя сделать никаких выводов. Совсем недавно была оценена роль трансплантации стволовых клеток (SCT) у пациентов с PBL. 54 Похоже, что у пациентов с ЛПК с чувствительным к химиотерапии заболеванием аутологичная ТСК может принести пользу в первой ремиссии. В итальянском исследовании сообщалось, что у 5 ВИЧ-позитивных пациентов с ЛПК, получавших высокодозную химиотерапию (ГДХ) с последующей аутологичной ТСК в первой ремиссии, удалось увеличить время общей выживаемости. 41 Аналогичным образом, серия случаев из 9 ВИЧ-отрицательных пациентов с ЛБЛ показала обнадеживающие результаты с 5-летней выживаемостью 60%. 10 В этом исследовании 4 пациентам была выполнена HDC с последующей аутологичной SCT в первой ремиссии. Опыт применения ГДК с последующей аутологичной ТСК при рецидивах довольно ограничен, хотя есть некоторые предположения, что при чувствительном к химиотерапии заболевании можно достичь стойкой полной ремиссии.
Лучевая терапия использовалась как часть лечения примерно в 20–30 случаях ЛБЛ, но на основании этого ограниченного опыта нельзя сделать никаких выводов. Совсем недавно была оценена роль трансплантации стволовых клеток (SCT) у пациентов с PBL. 54 Похоже, что у пациентов с ЛПК с чувствительным к химиотерапии заболеванием аутологичная ТСК может принести пользу в первой ремиссии. В итальянском исследовании сообщалось, что у 5 ВИЧ-позитивных пациентов с ЛПК, получавших высокодозную химиотерапию (ГДХ) с последующей аутологичной ТСК в первой ремиссии, удалось увеличить время общей выживаемости. 41 Аналогичным образом, серия случаев из 9 ВИЧ-отрицательных пациентов с ЛБЛ показала обнадеживающие результаты с 5-летней выживаемостью 60%. 10 В этом исследовании 4 пациентам была выполнена HDC с последующей аутологичной SCT в первой ремиссии. Опыт применения ГДК с последующей аутологичной ТСК при рецидивах довольно ограничен, хотя есть некоторые предположения, что при чувствительном к химиотерапии заболевании можно достичь стойкой полной ремиссии. 41 Применение аллогенной ТСК при ВИЧ-позитивных лейкоцитах показало ограниченную эффективность. 54
41 Применение аллогенной ТСК при ВИЧ-позитивных лейкоцитах показало ограниченную эффективность. 54
Учитывая плохие результаты и выживаемость пациентов с ЛБЛ, необходимы новые агенты. Были предприняты попытки использования антимиеломных агентов на основе плазмоцитарной дифференцировки клеток PBL, хотя опыт ограничен сообщениями о случаях заболевания. Было показано, что ингибитор протеасом бортезомиб эффективен у пациентов с ДВККЛ после ГЦ, вызывая более высокие показатели ответа и выживаемости при использовании в сочетании со схемами, содержащими антрациклины. 55 Бортезомиб отдельно и в комбинации с химиотерапией использовался с ограниченной эффективностью у ВИЧ-положительных и ВИЧ-отрицательных пациентов с рецидивом ЛБЛ. 56-62 Совсем недавно в серии случаев у 3 ранее не леченных пациентов с ЛБЛ, 2 из которых были ВИЧ-позитивными, была показана эффективность комбинации бортезомиба и EPOCH с поправкой на дозу.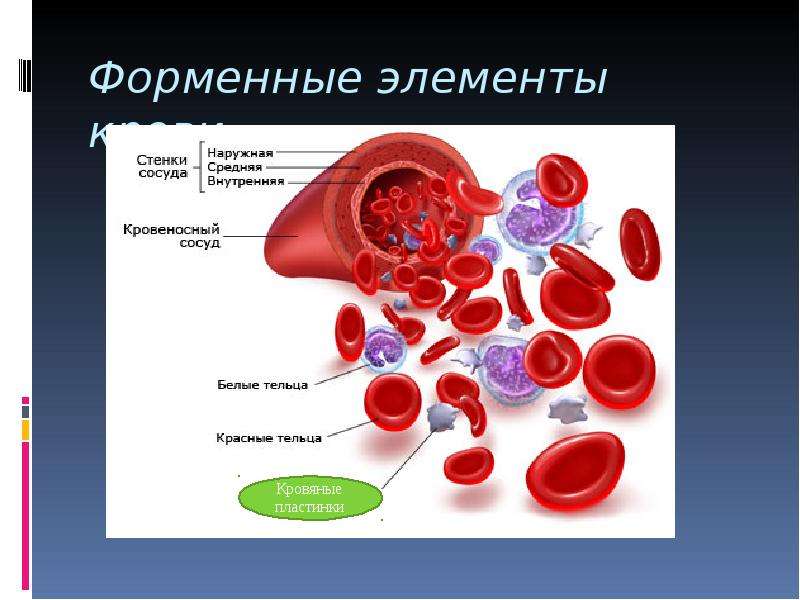 63 Иммуномодулятор леналидомид вызывал временный ответ у пациента с рецидивом ЛБЛ. 56 Исследования показали, что примерно в 30% случаев PBL экспрессируется маркер активации CD30, 4,28,64 , а недавний отчет показал ответ на брентуксимаб ведотин у пациента с CD30-экспрессирующим рецидивом PBL. 65 Однако специфическое пороговое значение CD30-позитивности среди клеток лимфомы не определено.
63 Иммуномодулятор леналидомид вызывал временный ответ у пациента с рецидивом ЛБЛ. 56 Исследования показали, что примерно в 30% случаев PBL экспрессируется маркер активации CD30, 4,28,64 , а недавний отчет показал ответ на брентуксимаб ведотин у пациента с CD30-экспрессирующим рецидивом PBL. 65 Однако специфическое пороговое значение CD30-позитивности среди клеток лимфомы не определено.
Наша рекомендация для лечения ЛБЛ первой линии — 6 циклов инфузионных дозозависимых EPOCH (с бортезомибом или без него) с интратекальной профилактикой в каждом цикле EPOCH и рассмотрением возможности консолидирующей HDC с последующей аутологичной ТСК в первой ремиссии для соответствующих кандидатов. У ВИЧ-позитивных пациентов кАРТ следует начинать или оптимизировать под наблюдением специалиста по инфекционным заболеваниям, имеющего опыт изучения возможных взаимодействий между противоопухолевыми препаратами и кАРТ. Мы рекомендуем лучевую терапию в паллиативных условиях, хотя ее можно рассматривать как консолидацию в каждом конкретном случае после проведения полного курса комбинированной химиотерапии.
Мы рекомендуем лучевую терапию в паллиативных условиях, хотя ее можно рассматривать как консолидацию в каждом конкретном случае после проведения полного курса комбинированной химиотерапии.
Кровь и содержащиеся в ней клетки – группы крови и антигены эритроцитов
У среднего взрослого человека более 5 литров (6 кварт) крови в тело. Кровь переносит кислород и питательные вещества к живым клеткам и забирает их отходы продукты. Он также доставляет иммунные клетки для борьбы с инфекциями и содержит тромбоциты. которые могут образовывать пробку в поврежденном кровеносном сосуде, чтобы предотвратить потерю крови.
Через систему кровообращения кровь адаптируется к потребностям организма. Когда вы тренируясь, ваше сердце качается сильнее и быстрее, чтобы обеспечить больше крови и, следовательно, кислород к вашим мышцам. Во время инфекции кровь доставляет больше иммунных клеток к место заражения, где они накапливаются для защиты от вредных захватчиков.
Все эти функции делают кровь драгоценной жидкостью. Ежегодно в США 30 млн.
ед. компонентов крови переливают нуждающимся в них больным. Кровь считается
такое драгоценное, что его еще называют «красным золотом», потому что клетки и белки его
содержит, может быть продан дороже, чем стоимость того же веса в золоте.
Ежегодно в США 30 млн.
ед. компонентов крови переливают нуждающимся в них больным. Кровь считается
такое драгоценное, что его еще называют «красным золотом», потому что клетки и белки его
содержит, может быть продан дороже, чем стоимость того же веса в золоте.
В этой главе представлены компоненты крови.
Кровь содержит клетки, белки и сахара
Если пробирку с кровью оставить для выдерживают полчаса, кровь разделяется на три слоя по мере того, как более плотная компоненты опускаются на дно трубки, а жидкость остается наверху.Жидкость соломенного цвета, образующая верхний слой, называется плазмой и составляет около 60% крови. Средний белый слой состоит из лейкоцитов (лейкоцитов) и тромбоциты, а нижний красный слой — эритроциты (эритроциты).Эти два нижних слои клеток составляют около 40% крови.
Плазма в основном состоит из воды, но она также содержит много важных веществ, таких как
белки (альбумины, факторы свертывания крови, антитела, ферменты и гормоны), сахара
(глюкоза) и частицы жира.
Все клетки крови происходят из костного мозга. Они начинают свою жизнь как стволовые клетки, и они созревают в три основных типа клеток — эритроциты, лейкоциты, и тромбоциты. В свою очередь различают три типа лейкоцитов — лимфоциты, моноциты и гранулоциты – и три основных типа гранулоцитов (нейтрофилы, эозинофилы и базофилы).Посмотрите их в действии в разделе «Знакомство с клетками крови».
См. рисунок всех клеточных элементов крови в журнале Janeway & Traver’s Immunobiology.
Образец крови может быть дополнительно разделен на отдельные компоненты путем центрифугирования
образец в центрифуге. Сила вращения заставляет более плотные элементы
сток, а дальнейшая обработка позволяет выделить конкретный белок или
выделение определенного типа клеток крови. С использованием этого метода,
антитела и факторы свертывания могут быть получены из плазмы для лечения иммунных
недостаточность и нарушение свертываемости соответственно.Точно так же можно собирать эритроциты.
для переливания крови.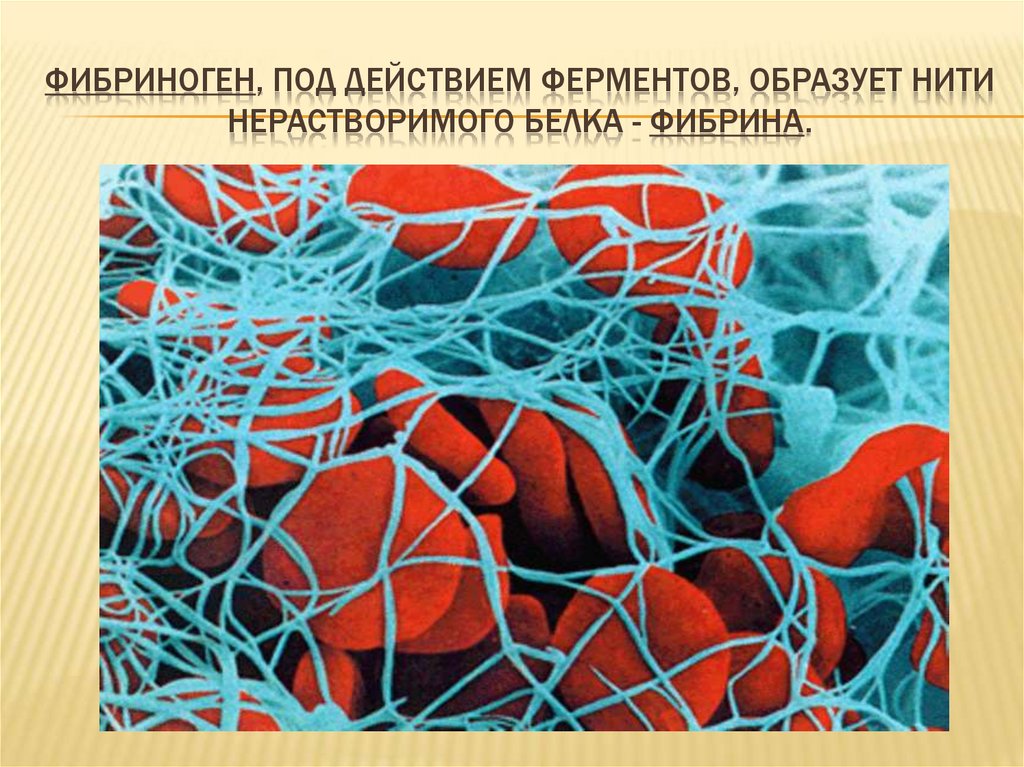
Красные кровяные тельца переносят кислород
Каждую секунду 2-3 миллиона эритроцитов вырабатывается в костном мозге и высвобождается в кровоток. Также известен как эритроциты, эритроциты являются наиболее распространенным типом клеток, обнаруживаемых в крови, причем каждый кубический миллиметр крови, содержащей 4-6 миллионов клеток. Диаметром всего 6 мкм, эритроциты достаточно малы, чтобы протискиваться через мельчайшие кровеносные сосуды. Они циркулируют по телу до 120 дней, после чего старые или поврежденные Эритроциты выводятся из кровотока специализированными клетками (макрофагами) в селезенка и печень. У человека, как и у всех млекопитающих, в зрелых эритроцитах отсутствует ядро. Это позволяет клетке
больше места для хранения гемоглобина, белка, связывающего кислород, что позволяет эритроцитам
транспортировать больше кислорода. Эритроциты также имеют двояковогнутую форму; эта форма увеличивает их
площадь поверхности для диффузии кислорода через их поверхности. У не млекопитающих
у позвоночных, таких как птицы и рыбы, зрелые эритроциты имеют ядро.
Если у пациента низкий уровень гемоглобина, состояние, называемое анемией, он может кажутся бледными, потому что гемоглобин придает эритроцитам, а следовательно, и крови их красный цвет.Они также может легко утомляться и чувствовать одышку из-за важной роли гемоглобина в транспортировке кислорода из легких туда, где он необходим. тело.
Лейкоциты являются частью иммунного ответа
Лейкоциты бывают разных форм и размеров. Некоторые клетки имеют ядра с несколькими доли, тогда как другие содержат одно большое круглое ядро. Некоторые содержат пакеты гранулы в их цитоплазме и поэтому известны как гранулоциты.
Несмотря на различия во внешнем виде, все различные типы лейкоцитов играют определенную роль.
в иммунном ответе.Они циркулируют в крови до тех пор, пока не получат сигнал о том, что
часть тела повреждена. Сигналы включают интерлейкин 1 (IL-1), молекулу
секретируется макрофагами, что способствует лихорадке при инфекциях, и гистамином,
высвобождается циркулирующими базофилами и тучными клетками тканей и способствует
аллергические реакции. В ответ на эти сигналы лейкоциты покидают кровеносный сосуд
сдавливание через отверстия в стенке кровеносного сосуда. Они мигрируют к источнику
сигнализировать и помочь начать процесс заживления.
В ответ на эти сигналы лейкоциты покидают кровеносный сосуд
сдавливание через отверстия в стенке кровеносного сосуда. Они мигрируют к источнику
сигнализировать и помочь начать процесс заживления.
Лица с низким уровнем лейкоцитов могут иметь более тяжелые инфекции. В зависимости при отсутствии лейкоцитов пациент подвергается риску различных типов инфекционное заболевание. Например, макрофаги особенно хорошо поглощают бактерии. дефицит макрофагов приводит к рецидивирующим бактериальным инфекциям. Напротив, Т клетки особенно хорошо борются с вирусными инфекциями, и потеря их Функция приводит к повышенной восприимчивости к вирусным инфекциям.
Нейтрофилы переваривают бактерии
Нейтрофилы также известны как полиморфноядерные клетки, потому что они содержат ядро, форма (морф) которого неправильной формы и содержит много (поли) долей.Они также принадлежат к группе лейкоцитов. называют гранулоцитами, потому что их цитоплазма усеяна гранулами, содержат ферменты, помогающие им переваривать болезнетворные микроорганизмы.
Моноциты становятся макрофагами
Моноциты представляют собой молодые лейкоциты, циркулировать в крови. Они превращаются в макрофаги после того, как покидают кровь и мигрируют в ткани. Там они обеспечивают немедленную защиту, потому что они могут поглощать (фагоцитировать) и переваривать патогены раньше других типов лейкоцитов. добраться до района.В печени тканевые макрофаги называются клетками Купфера, и они специализируются на удаление вредных агентов из крови, вышедшей из кишечника.Альвеолярные макрофаги находятся в легких и удаляют вредные вещества, которые могли попасть в дыхательные пути. Макрофаги в селезенке удаляют старые или поврежденные эритроциты и тромбоциты. из тиража.
Макрофаги также являются «антигенпрезентирующими клетками», представляющими чужеродные белки (антигены) к другим иммунным клеткам, вызывая иммунный ответ.
Лимфоциты состоят из В-клеток и Т-клеток
Лимфоциты представляют собой круглые клетки, содержат одно крупное круглое ядро. Различают два основных класса клеток,
В-клетки, созревающие в костном мозге, и Т-клетки, созревающие в
вилочковая железа.
Различают два основных класса клеток,
В-клетки, созревающие в костном мозге, и Т-клетки, созревающие в
вилочковая железа.После активации В-клетки и Т-клетки запускают различные типы иммунных отклик. Активированные В-клетки, также известные как плазматические клетки, производят большое количество специфические антитела, которые связываются с агентом, вызвавшим иммунный ответ. Т клетки, называемые вспомогательными Т-клетками, выделяют химические вещества, которые рекрутируют другие иммунные клетки. и помогите скоординировать их атаку. Другая группа, называемая цитотоксическими Т-клетками, атакует инфицированные вирусом клетки.
Тромбоциты способствуют свертыванию крови
Тромбоциты неправильной формы фрагменты клеток, которые циркулируют в крови до тех пор, пока они либо не активируются для образуют кровяной сгусток или удаляются селезенкой.Тромбоцитопения – это состояние низкий уровень тромбоцитов и несет повышенный риск кровотечения. И наоборот, высокий уровень тромбоцитов (тромбоцитемия) несет в себе повышенный риск формирования несоответствующие кровяные сгустки.Как и все клетки крови, тромбоциты происходят из стволовых клеток в костях. костный мозг. Стволовые клетки превращаются в предшественники тромбоцитов (мегакариоциты), которые «сбросить» тромбоциты в кровь.Там тромбоциты циркулируют около 9 дней. Если в это время они сталкиваются с поврежденными стенками кровеносных сосудов, они прилипают к поврежденном участке и активируются, образуя кровяной сгусток. Это затыкает дыру. В противном случае, в конце жизни они удаляются из кровообращения селезенкой. При различных заболеваниях, при которых селезенка гиперактивна, напр. ревматоидный артрите и лейкемии, селезенка удаляет слишком много тромбоцитов, что приводит к увеличению кровотечение.
Ваш общий анализ крови
Общий анализ крови (CBC) — это простой анализ крови, который обычно заказывают как часть
обычного медицинского освидетельствования.Как следует из названия, это счет
различных типов клеток, обнаруженных в крови. Тест может диагностировать и контролировать многие
различные заболевания, такие как анемия, инфекции, воспалительные заболевания и
злокачественность. дает пример
значения CBC, но обратите внимание, что референтные диапазоны и используемые единицы измерения могут различаться.
в зависимости от лаборатории, проводившей исследование.
Тест может диагностировать и контролировать многие
различные заболевания, такие как анемия, инфекции, воспалительные заболевания и
злокачественность. дает пример
значения CBC, но обратите внимание, что референтные диапазоны и используемые единицы измерения могут различаться.
в зависимости от лаборатории, проводившей исследование.
Подсчет эритроцитов позволяет выявить анемию
ОАК измеряет следующие характеристики эритроцитов:
общее количество гемоглобина (Hb) в крови средний размер эритроцитов (MCV)
количество места, которое эритроциты занимают в крови (гематокрит)
Общий анализ крови также включает информацию о эритроцитах, которая рассчитывается из других измерения, т.г., количества (МСН) и концентрации (МСНС) гемоглобина в эритроциты.
Количество эритроцитов и количество гемоглобина в крови ниже у женщин
чем у мужчин. Это происходит из-за менструальной потери крови каждый месяц. Ниже
определенного уровня гемоглобина, говорят, что у пациента анемия, что предполагает
клинически значимое снижение кислородной емкости. Анемия не является
диагноз, а симптом основного заболевания, которое необходимо исследовать.
Анемия не является
диагноз, а симптом основного заболевания, которое необходимо исследовать.
Ключом к разгадке причины анемии является средний размер эритроцитов (средний корпускулярный объем, MCV).Причины высокого MCV включают дефицит B 12 или витамины фолиевой кислоты в рационе. B 12 содержится в красном мясе, поэтому дефицит B 12 особенно часто встречается у вегетарианцев и веганов. И наоборот, фолиевой кислоты много в свежих листовых зеленых овощах, поэтому Дефицит фолиевой кислоты часто встречается у пожилых людей, которые могут плохо питаться.
Анемия с низким MCV встречается часто и может быть результатом наследственных заболеваний крови, таких как как талассемия, но чаще всего вызвана дефицитом железа.Например, женщины репродуктивного возраста могут терять слишком много железа из-за обильных менструаций кровотечения и склонны к этой форме анемии, известной как железодефицитная анемия.
Гематокрит — это процент эритроцитов по отношению к общему объему крови.
Гематокрит измеряет долю крови, состоящую из эритроцитов. Это отражает сочетание общего количества эритроцитов и объема, который они занимать.
Одним из изменений, наблюдаемых во время беременности, является падение гематокрита.Это происходит потому, что хотя продукция эритроцитов существенно не меняется, объем плазмы увеличивается, т. е. эритроциты «разбавляются». Кроме того, низкий гематокрит может отражают снижение продукции эритроцитов костным мозгом. Это может быть связано с заболевания костного мозга (повреждение токсинами или раком) или из-за снижения эритропоэтин, гормон, секретируемый почками, который стимулирует выработку эритроцитов. Снижение числа эритроцитов также может быть результатом сокращения продолжительности жизни эритроцитов (например, хроническое кровотечение).
Высокое значение гематокрита может действительно отражать увеличение доли эритроцитов
(например, повышенный уровень эритропоэтина, связанный с опухолью эритроцитов, называемой
красная полицитемия), или это может отражать снижение плазменного компонента
кровь (например, потеря жидкости у пострадавших от ожогов).
Увеличение числа лейкоцитов при инфекциях и опухолях
Количество лейкоцитов — это количество лейкоцитов, обнаруженных в кровь.
Повышенное количество лейкоцитов чаще всего вызывается инфекциями, такими как инфекция мочевыводящих путей или пневмония.Это также может быть вызвано опухолями WBC, такими как как лейкемия.
Снижение количества лейкоцитов вызвано тем, что костный мозг не может производить лейкоциты или повышенным удалением лейкоцитов из кровотока больной печенью или гиперактивная селезенка. Отказ костного мозга может быть вызван токсинами или нормальные клетки костного мозга замещаются опухолевыми клетками.
Дифференциальная часть WBC CBC разбивает WBC на пять различных
типы: нейтрофилы, лимфоциты, моноциты, эозинофилы и базофилы.Находка
количество лейкоцитов каждого типа дает больше информации о лежащем в их основе
проблема. Например, на ранних стадиях инфекции большая часть увеличения
в лейкоцитах объясняется увеличением нейтрофилов.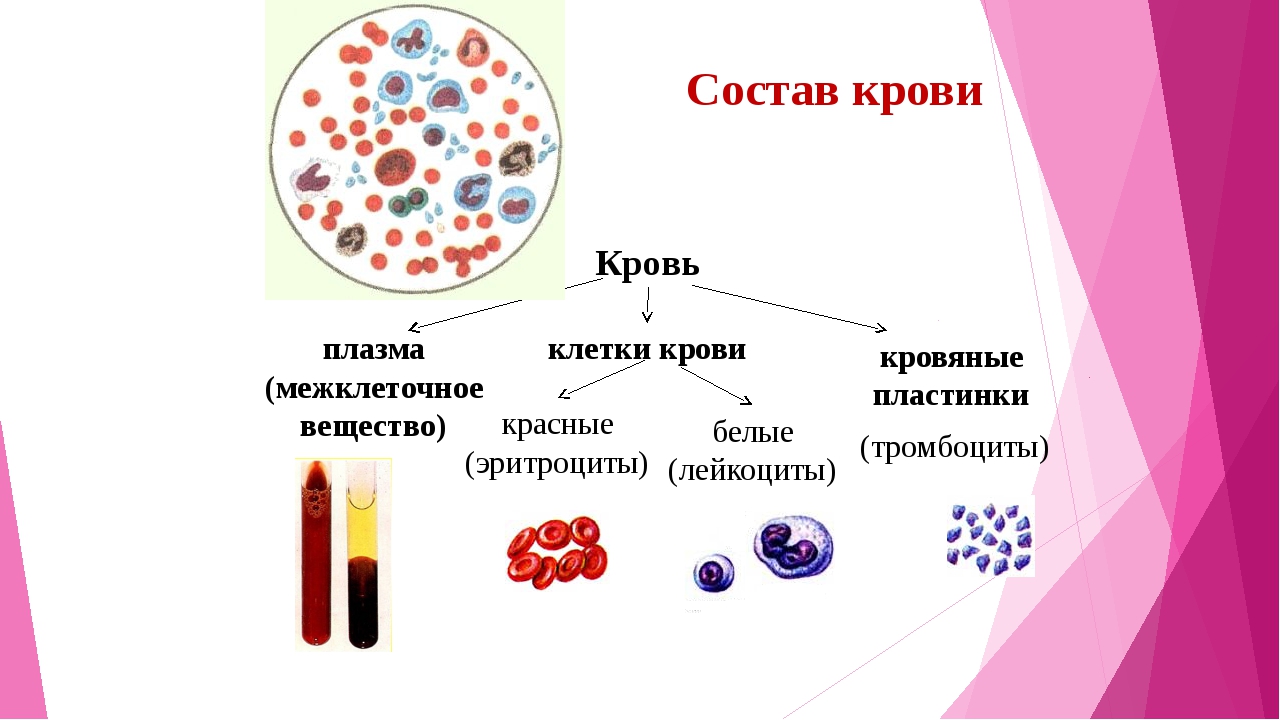 Как инфекция
продолжается, лимфоциты увеличиваются. Заражение гельминтами может вызвать увеличение
эозинофилов, в то время как аллергические состояния, такие как сенная лихорадка, вызывают увеличение
в базофилах.
Как инфекция
продолжается, лимфоциты увеличиваются. Заражение гельминтами может вызвать увеличение
эозинофилов, в то время как аллергические состояния, такие как сенная лихорадка, вызывают увеличение
в базофилах.
Количество тромбоцитов указывает на вероятность кровотечения или свертывания
В норме один кубический миллиметр крови содержит от 150 000 до 400 000 тромбоциты.Если число падает ниже этого диапазона, неконтролируемое кровотечение становится риск, тогда как превышение верхней границы этого диапазона указывает на риск неконтролируемое свертывание крови.
Гемоглобин связывает кислород
Гемоглобин — белок, переносящий кислород, который содержится во всех эритроцитах. Он поднимает кислород там, где его много (легкие), и выбрасывает кислород там, где он необходим вокруг тела. Гемоглобин также является пигментом, придающим эритроцитам красный цвет.
Гемовые группы и глобины
Как следует из названия, гемоглобин состоит из «гемовых» групп (железосодержащих
кольца) и «глобины» (белки). На самом деле гемоглобин состоит из четырех глобинов.
белки — две альфа-цепи и две бета-цепи — каждая с
гемовая группа. Группа гема содержит один атом железа, и он может связывать один
молекула кислорода. Поскольку каждая молекула гемоглобина содержит четыре глобина,
он может переносить до четырех молекул кислорода.
На самом деле гемоглобин состоит из четырех глобинов.
белки — две альфа-цепи и две бета-цепи — каждая с
гемовая группа. Группа гема содержит один атом железа, и он может связывать один
молекула кислорода. Поскольку каждая молекула гемоглобина содержит четыре глобина,
он может переносить до четырех молекул кислорода.
Гемоглобин переносит кислород
В легких молекула гемоглобина окружена высокой концентрацией кислород, следовательно, он связывает кислород. В активных тканях концентрация кислорода ниже, поэтому гемоглобин выделяет кислород.
Такое поведение намного эффективнее, потому что Связывание гемоглобина с кислородом является «кооперативным». Это означает что связывание одной молекулы кислорода облегчает связывание последующие молекулы кислорода. Точно так же освобождение кислорода облегчает для высвобождения других молекул кислорода. Это означает, что реакция гемоглобина к кислородным потребностям активных тканей намного быстрее.
Помимо насыщения гемоглобина кислородом, другие факторы, влияющие на
готовность гемоглобина связывать кислород, включая pH плазмы, уровень бикарбоната плазмы,
и давление кислорода в воздухе (особенно на больших высотах).
Молекула 2,3-дифосфоглицерат (2,3-ДФГ) связывается с гемоглобином и снижает его сродство к кислороду, что способствует выделению кислорода. У лиц, имеющих акклиматизировались к жизни на больших высотах, уровень 2,3-ДФГ в крови увеличивается, что позволяет доставлять больше кислорода к тканям при низком уровне напряжение кислорода.
Фетальный гемоглобин
Фетальный гемоглобин отличается от взрослого гемоглобина тем, что содержит два гамма цепочки вместо двух бета-цепочек. Фетальный гемоглобин связывает кислород с большим большее сродство, чем у взрослого гемоглобина; это преимущество в утробе матери, потому что он позволяет крови плода извлекать кислород из материнской крови, несмотря на его низкую концентрация кислорода.
В норме весь фетальный гемоглобин замещается взрослым гемоглобином ко времени рождение.
Расщепление гемоглобина
Старые или поврежденные эритроциты удаляются из кровотока макрофагами в селезенке
и печени, а содержащийся в них гемоглобин расщепляется на гем и глобин.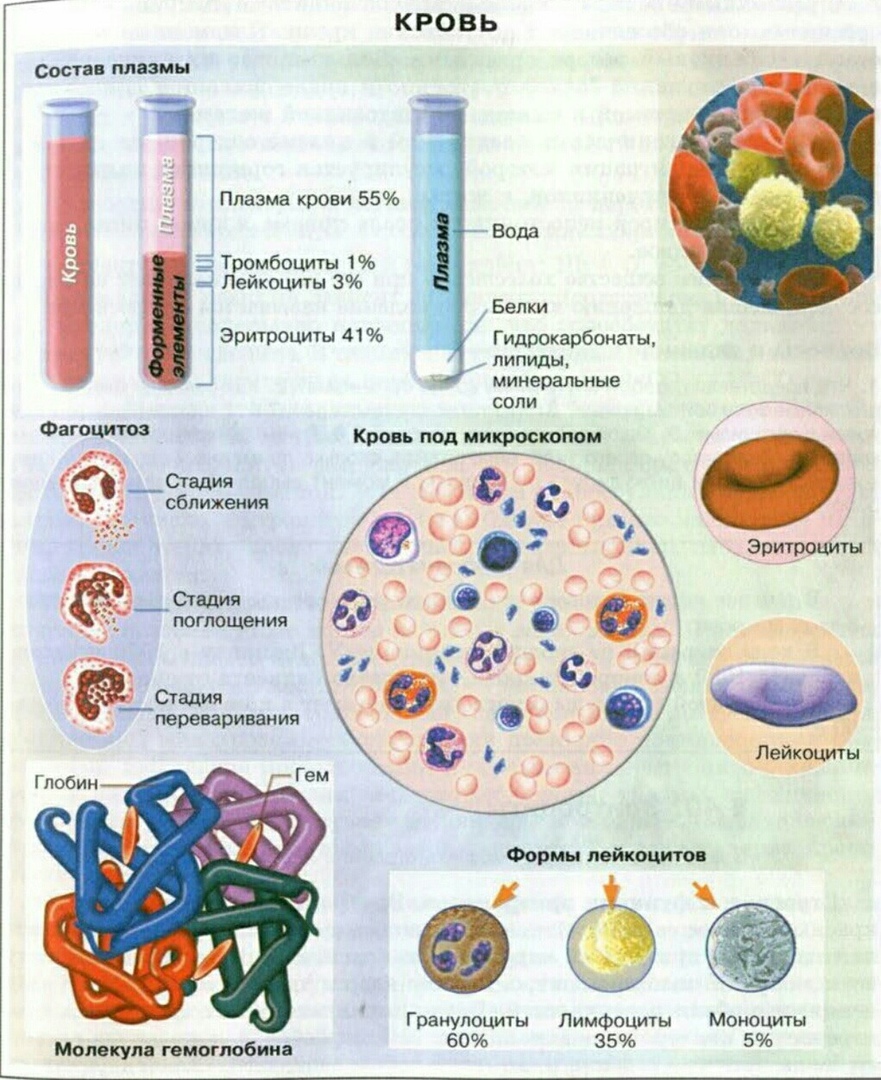 Белок глобина может быть переработан или расщеплен до его составляющих.
аминокислоты, которые могут быть переработаны или метаболизированы. Гем содержит драгоценные
железо, которое сохраняется и повторно используется в синтезе новых молекул гемоглобина.
Белок глобина может быть переработан или расщеплен до его составляющих.
аминокислоты, которые могут быть переработаны или метаболизированы. Гем содержит драгоценные
железо, которое сохраняется и повторно используется в синтезе новых молекул гемоглобина.
В процессе метаболизма гем превращается в билирубин, желтый пигмент, который может обесцвечивать кожу и склеру глаза, если он накапливается в крови, состояние, известное как желтуха. Вместо этого белок плазмы альбумин связывается с билирубина и переносит его в печень, где он выделяется с желчью, а также способствует окраске фекалий.
Желтуха – одно из осложнений переливания несовместимой крови. Этот возникает, когда иммунная система реципиента атакует донорские эритроциты как иностранный.Скорость разрушения эритроцитов и последующего образования билирубина может превышают способность печени метаболизировать вырабатываемый билирубин.
Гемоглобинопатии
Гемоглобинопатии образуют группу наследственных заболеваний, вызываемых
мутации в глобиновых цепях гемоглобина. Серповидноклеточная анемия является наиболее
распространены среди них и связаны с мутацией, которая изменяет одну из аминогрупп
кислоты в бета-цепи гемоглобина, образуя «хрупкий» гемоглобин. Когда
концентрация кислорода низкая, эритроциты имеют тенденцию к деформации и серповидности.
в форме.Эти деформированные клетки могут блокировать мелкие кровеносные сосуды и повреждать органы.
они поставляют. Это может быть очень болезненно, и если не лечить, серповидно-клеточная анемия
кризис может быть фатальным.
Серповидноклеточная анемия является наиболее
распространены среди них и связаны с мутацией, которая изменяет одну из аминогрупп
кислоты в бета-цепи гемоглобина, образуя «хрупкий» гемоглобин. Когда
концентрация кислорода низкая, эритроциты имеют тенденцию к деформации и серповидности.
в форме.Эти деформированные клетки могут блокировать мелкие кровеносные сосуды и повреждать органы.
они поставляют. Это может быть очень болезненно, и если не лечить, серповидно-клеточная анемия
кризис может быть фатальным.
Еще одна наследственная анемия, особенно поражающая жителей Средиземноморья происхождение – талассемия. Нарушение выработки альфа- или бета-глобина цепи вызывает ряд симптомов, в зависимости от того, сколько копий альфа и затронуты бета-гены. Некоторые люди могут быть переносчиками заболевания и не имеют симптомов, тогда как если все копии генов потеряны, болезнь фатальный.
Порфирии представляют собой группу наследственных заболеваний, при которых синтез гема
нарушен. В зависимости от стадии, на которой происходит нарушение, различают
ряд неврологических и желудочно-кишечных побочных эффектов. король Георг III
Англия («Безумие короля Георга») была одной из самых известных личностей
кто страдал порфирией.
король Георг III
Англия («Безумие короля Георга») была одной из самых известных личностей
кто страдал порфирией.
Все дело в равновесии · Границы для юных умов
Аннотация
Независимо от того, являетесь ли вы человеком, летучей мышью или садовым кустарником, вам нужно железо, чтобы выжить.Железо необходимо для самых фундаментальных процессов жизни. Однако в избытке он очень токсичен для организма. В результате, когда в вашем организме слишком мало или слишком много железа, вы можете сильно заболеть. В этой статье мы обсудим, как мы получаем достаточное количество железа и куда оно уходит в нашем организме после употребления. Вы также узнаете о болезнях, связанных с железом, в том числе о заболевании летучих мышей в зоопарке в 1980-х годах.
Больные летучие мыши в зоопарке: История железа
Около 30 лет назад в зоопарке Торонто серьезно заболел вид летучих мышей, называемых египетскими фруктовыми летучими мышами ( Rousettus egyptiacus ). Эти фруктовые летучие мыши настолько заболели, что некоторые из них даже умерли. Примерно в то же время у летучих мышей в других зоопарках также проявлялись аналогичные симптомы. Что могло послужить причиной того, что фруктовые летучие мыши так заболели? По мере того, как ученые пытались найти причину, они начали исключать определенные причины. Например, они не думали, что болезнь была вызвана инфекционным заболеванием, и не думали, что эти летучие мыши умерли от естественных причин, поскольку они были разного возраста и были здоровы до начала болезни.
Эти фруктовые летучие мыши настолько заболели, что некоторые из них даже умерли. Примерно в то же время у летучих мышей в других зоопарках также проявлялись аналогичные симптомы. Что могло послужить причиной того, что фруктовые летучие мыши так заболели? По мере того, как ученые пытались найти причину, они начали исключать определенные причины. Например, они не думали, что болезнь была вызвана инфекционным заболеванием, и не думали, что эти летучие мыши умерли от естественных причин, поскольку они были разного возраста и были здоровы до начала болезни.
Потенциальная подсказка связана с их диетой.В зоопарке Торонто летучих мышей в основном кормили мягкими фруктами, которыми также питаются египетские фруктовые летучие мыши, живущие в дикой природе в Африке к югу от Сахары, в Северной Африке и на Ближнем Востоке [2]. Однако смотрители зоопарка добавляли в пищу летучих мышей порошок, содержащий различные питательные вещества, похожие на жевательные резинки с витаминами, которые можно было принимать (рис. 1). Работники зоопарка давали летучим мышам этот порошок, чтобы они были здоровы, но один из компонентов, железо, мог непреднамеренно вызвать у них заболевание. В частности, в порошке было слишком много железа по сравнению с количеством железа, которое фруктовые летучие мыши обычно едят в дикой природе.
1). Работники зоопарка давали летучим мышам этот порошок, чтобы они были здоровы, но один из компонентов, железо, мог непреднамеренно вызвать у них заболевание. В частности, в порошке было слишком много железа по сравнению с количеством железа, которое фруктовые летучие мыши обычно едят в дикой природе.
- Рис. 1. Рацион египетских фруктовых летучих мышей в дикой природе и в зоопарке.
- (A) Египетские фруктовые летучие мыши обычно питаются (B) мягкими фруктами, но в зоопарке (C) к их корму добавляли порошок, содержащий железо и другие питательные вещества.
Любопытно, что другой вид летучих мышей в зоопарке, летучая мышь-вампир, не заболел, несмотря на то, что потреблял пищу, богатую железом. Чтобы лучше понять, почему избыток железа может вызывать болезни у фруктовых летучих мышей и при этом щадить летучих мышей-вампиров, сначала необходимо понять, что такое железо и почему оно важно в умеренных количествах для благополучия животных.
Важность железа
Железо является элементом, необходимым для большинства живых организмов на Земле. Деревья за вашим окном нуждаются в железе, чтобы выжить так же, как вы, ваша собака или бактерии, живущие вокруг вас. Животные обычно получают железо, употребляя в пищу железосодержащие продукты. Возможно, вы уже слышали, что красное мясо богато железом, но железо также содержится в яйцах, морепродуктах, шпинате, тофу и множестве других продуктов. Иногда пища может не содержать железа естественным образом, но железо добавляется в нее при обработке, потому что это необходимое питательное вещество для поддержания здоровья людей.В результате даже ваше молоко или хлопья для завтрака могут содержать железо.
Почему железо так важно? Это ключевой компонент многих фундаментальных процессов, необходимых для выживания. Например, он нужен нам для создания энергии для подпитки нашего тела, а также для правильного использования кислорода, которым мы дышим [3]. Однако существует тонкий баланс, потому что слишком много железа также может нанести нам серьезный ущерб. В результате наши тела нашли способы регулировать количество железа внутри нас в любой момент времени.
В результате наши тела нашли способы регулировать количество железа внутри нас в любой момент времени.
Путешествие железа по телу
Вам интересно, как наш организм усваивает и использует железо? Чтобы это выяснить, давайте проследим путь железа по всему человеческому телу. Хотя мы будем использовать пример человека, аналогичные процессы происходят и у других животных, например у летучих мышей.
Когда вы едите железосодержащую пищу, такую как красное мясо, она сначала попадает в желудок и начинает перевариваться (рис. 2). Затем в тонком кишечнике, который является частью пищеварительной системы, которая соединяется с желудком, железо всасывается.Для этого поглощения существует специальная структура, называемая переносчиком железа , которая позволяет железу перемещаться между клетками организма. Железо нуждается в переносчиках железа для перемещения, потому что каждая клетка окружена мембраной, которая является самым внешним слоем клетки. Думайте о мембране как о стенах, окружающих вашу комнату. Вместо того, чтобы швыряться о стену в надежде попасть в следующую комнату, вы будете использовать двери, чтобы попасть между комнатами, что похоже на то, как железо использует переносчики железа для перемещения от клетки к клетке внутри тела [4].
Вместо того, чтобы швыряться о стену в надежде попасть в следующую комнату, вы будете использовать двери, чтобы попасть между комнатами, что похоже на то, как железо использует переносчики железа для перемещения от клетки к клетке внутри тела [4].
- Рисунок 2. Путь железа через тело человека.
- (A) Строение желудка, кишечника и печени человека. Железо всасывается через слизистую оболочку тонкой кишки с помощью переносчиков железа. (B) Структура кости человека с изображением костного мозга и крупным планом эритроцита.
Когда железо всасывается в пищеварительной системе, оно проходит через кровь, чтобы попасть в другие места по всему телу.Поскольку железо может быть токсичным, молекулы в крови, называемые белками, окружают его на протяжении всего процесса транспортировки, поэтому оно не может причинить нам вреда. Большая часть железа в нашем организме поступает в костный мозг, ткань в центре наших костей (рис. 2). Это связано с тем, что костный мозг вырабатывает эритроциты, которые переносят кислород по всему телу. Красная кровь содержит гемоглобина , который представляет собой белок в форме пончика, который нуждается в железе, чтобы переносить кислород в своем центре. Другими словами, большая часть железа в организме поступает в костный мозг, потому что гемоглобин нуждается в нем для переноса кислорода к остальным частям тела.
2). Это связано с тем, что костный мозг вырабатывает эритроциты, которые переносят кислород по всему телу. Красная кровь содержит гемоглобина , который представляет собой белок в форме пончика, который нуждается в железе, чтобы переносить кислород в своем центре. Другими словами, большая часть железа в организме поступает в костный мозг, потому что гемоглобин нуждается в нем для переноса кислорода к остальным частям тела.
Часть железа, которое мы едим, откладывается для дальнейшего использования. В частности, железо поступает в печень, орган в верхней правой части живота, расположенный под грудной клеткой (рис. 2). Печень выполняет множество функций, одна из которых – запасание железа.
Железо: все дело в балансе
Учитывая, сколько важных жизненных процессов зависит от железа, вы были бы правы, предположив, что без достаточного количества железа вы сильно заболеете. Это состояние называется железодефицитной анемией и может возникнуть по разным причинам, включая потерю крови и недостаток железа в вашем рационе. Железодефицитная анемия вызывает у вас слабость и усталость, и даже может вызвать желание есть лед или грязь. К счастью, существует относительно простое лекарство от этого состояния в виде таблеток железа, которые может назначить врач [3].
Железодефицитная анемия вызывает у вас слабость и усталость, и даже может вызвать желание есть лед или грязь. К счастью, существует относительно простое лекарство от этого состояния в виде таблеток железа, которые может назначить врач [3].
На противоположной стороне спектра перегрузка железом — это состояние, которое возникает, когда в организме слишком много железа. Пациенты с перегрузкой железом также могут испытывать такие симптомы, как слабость и утомляемость. Состояние может даже привести к повреждению органов, таких как печень, где хранится железо.Один из методов лечения перегрузки железом включает забор крови у пациентов до тех пор, пока уровень железа не снизится. Как железодефицитная анемия, так и перегрузка железом могут быть диагностированы врачом с помощью анализов крови.
К счастью для нас, у нас есть вещество под названием гепсидин , которое регулирует уровень железа в нашем организме (рис. 3) [3]. Когда в организме слишком много железа, количество гепсидина увеличивается, что приводит к снижению уровня железа.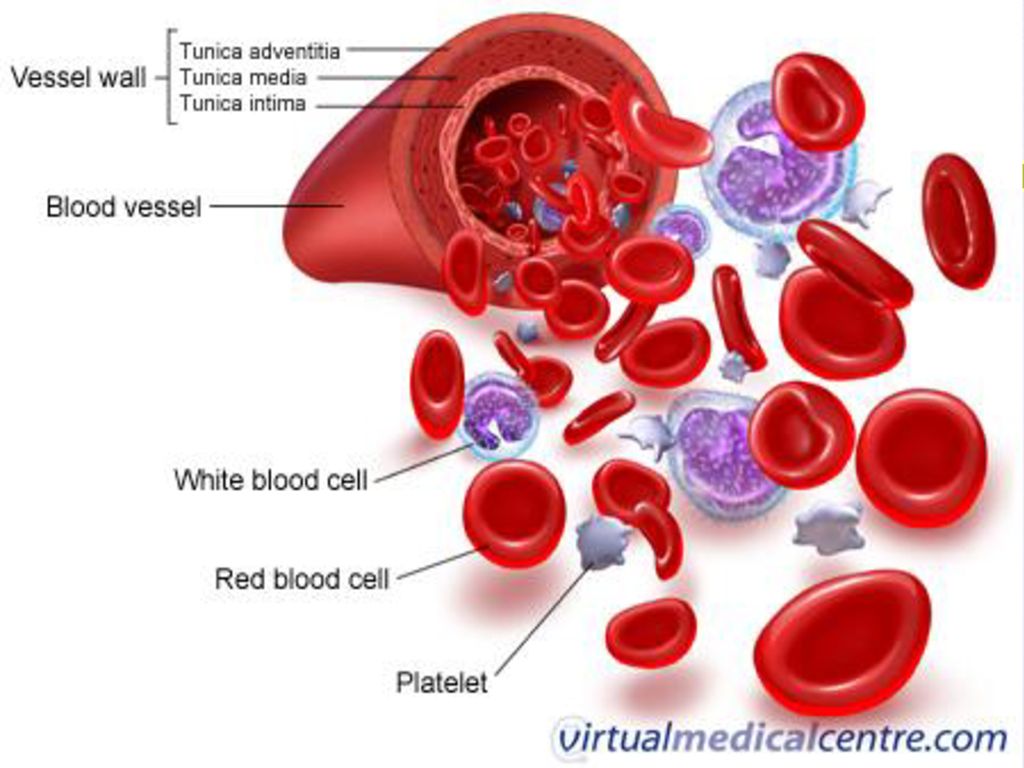 Когда железа не хватает, количество гепсидина уменьшается, что позволяет организму усваивать больше потребляемого железа.
Когда железа не хватает, количество гепсидина уменьшается, что позволяет организму усваивать больше потребляемого железа.
- Рисунок 3 – Значение гепсидина в регуляции содержания железа в организме.
- (A) В ответ на избыток железа в организме повышается уровень гепсидина, что способствует снижению уровня железа. (B) Когда железа слишком мало, уровень гепсидина уменьшается, чтобы позволить большему количеству железа попасть в организм.
Последняя часть головоломки: гепсидин
Вернемся к случаю с египетскими фруктовыми летучими мышами в зоопарке. Зная об опасности железодефицитной анемии, вы теперь можете понять, почему смотрители зоопарка захотели добавить в пищу питательный порошок, содержащий железо.Теперь вы также понимаете, как этот порошок привел к перегрузке железом, что в конечном итоге привело к тому, что летучие мыши заболели и повредили их печень. Люди также могут получить перегрузку железом, если они потребляют слишком много железа или если их организм неправильно обрабатывает количество железа, которое они потребляют.
Как упоминалось ранее, осталась одна очень любопытная часть головоломки. Несмотря на то, что летучие мыши-вампиры в зоопарке питались той же богатой железом пищей, что и египетские фруктовые летучие мыши, они совсем не болели! Причина может быть связана с гепсидином, регулятором, который контролирует, сколько железа может быть поглощено организмом.Летучие мыши-вампиры резко отрегулировали количество гепсидина в своем организме, чтобы противодействовать повышенному содержанию железа в их пище, но египетским фруктовым летучим мышам было трудно регулировать уровень гепсидина [5]. Поскольку летучие мыши-вампиры привыкли потреблять кровь, богатую железом, их тела, возможно, приспособились переносить большое количество железа. Фруктовые летучие мыши, с другой стороны, привыкли к диете с низким содержанием железа. Возможно, поэтому их гепсидин плохо реагировал на избыток железа и, как следствие, почему они так заболевали от богатой железом диеты.
Резюме
Железо необходимо для нашего благополучия, но в избытке оно также вредно. Из пищи мы получаем железо, которое проходит обширный процесс, чтобы попасть из желудка в костный мозг, кровь и печень. Поскольку железо необходимо для жизненно важных процессов, слишком мало железа может привести к состоянию, называемому анемией, а слишком много — к перегрузке железом. Именно это и произошло с египетскими фруктовыми летучими мышами в зоопарке Торонто! Таким образом, живые организмы, включая вас, меня и египетских фруктовых летучих мышей, должны употреблять железосодержащие продукты в умеренных количествах, чтобы оставаться здоровыми.
Из пищи мы получаем железо, которое проходит обширный процесс, чтобы попасть из желудка в костный мозг, кровь и печень. Поскольку железо необходимо для жизненно важных процессов, слишком мало железа может привести к состоянию, называемому анемией, а слишком много — к перегрузке железом. Именно это и произошло с египетскими фруктовыми летучими мышами в зоопарке Торонто! Таким образом, живые организмы, включая вас, меня и египетских фруктовых летучих мышей, должны употреблять железосодержащие продукты в умеренных количествах, чтобы оставаться здоровыми.
Глоссарий
Транспортеры железа : ↑ Структуры, позволяющие железу перемещаться между мембранами к другим клеткам по всему телу.
Гемоглобин : ↑ Белок, который переносит кислород и требует железа для своего функционирования. Это компонент эритроцитов.
Железодефицитная анемия : ↑ Состояние, вызванное недостатком железа в организме, которое, помимо прочих симптомов, приводит к усталости и слабости.
Перегрузка железом : ↑ Состояние, вызванное избытком железа в организме, которое может вызвать повреждение органов.
Гепсидин : ↑ Белок, который регулирует уровень железа в нашем организме. Когда железа слишком много, гепсидин вызывает его снижение, а когда его слишком мало, гепсидин вызывает увеличение железа.
Конфликт интересов
Авторы заявляют, что исследование проводилось при отсутствии каких-либо коммерческих или финансовых отношений, которые могли бы быть истолкованы как потенциальный конфликт интересов.
Ссылки
[1] ↑ Кроушоу, Г., Оярсун, С., Вальдес, Э., и Роуз, К. 1995. «Гемохроматоз (болезнь накопления железа) у летучих мышей», в Трудах Первой конференции по зоопаркам. и Wildlife Nutrition (Скарборо, OT: Консультативная группа по питанию AZA).
[2] ↑ Красный список МСОП видов, находящихся под угрозой исчезновения . Доступно в Интернете по адресу: https://www.iucnredlist.org/species/29730/22043105 (по состоянию на 19 июня 2020 г.).
Доступно в Интернете по адресу: https://www.iucnredlist.org/species/29730/22043105 (по состоянию на 19 июня 2020 г.).
[3] ↑ Аббаспур, Н., Hurrell, R. и Kelishadi, R. 2014. Обзор железа и его важности для здоровья человека. Дж. Рез. Мед. науч. 19: 164–74.
[4] ↑ Вальдвогель-Абрамовски, С., Вэбер, Г., Гасснер, К., Бузер, А., Фрей, Б.М., Фаврат, Б., и др. 2014. Физиология обмена железа. Трансфус. Мед. мать. 41: 213–21. дои: 10.1159/000362888
[5] ↑ Стасиак И. М., Смит Д. А., Ганц Т., Кроушоу Г. Дж., Хаммермюллер Дж. Д., Бьенцле Д. и соавт.2018. Болезнь накопления железа (гемохроматоз) и реакция гепсидина на нагрузку железом у двух видов крылоногих летучих мышей по сравнению с обыкновенной летучей мышью-вампиром. J. Комп. Физиол. Б Биохим. Сист. Окружающая среда. Физиол. 188: 683–94. doi: 10.1007/s00360-018-1155-4
Кровообращение — онлайн-учебник по биологии
Система кровообращения человека
Кровь
Кровь состоит из жидкости, плазмы, и клеток крови, таких как эритроциты (эритроциты), лейкоциты (лейкоциты) и тромбоциты (клетки фрагменты). Гематокрит – процент объема крови, занимаемый эритроцитами.
Гематокрит – процент объема крови, занимаемый эритроцитами.
Плазма
Состоит из ряда неорганических и органических веществ: питательных веществ, метаболических отходов и гормонов, растворенных в воде. Он также содержит следующее:
- билирубин (продукт распада гемоглобина)
- альбумин (синтезируется печенью)
- глобулин
- фибриноген – участвует в свертывании крови.
Плазма, из которой удалены все белки, называется сывороткой.
Клетки крови
(1) Эритроциты
Эритроциты переносят кислород и углекислый газ, связывая их с железом гемоглобина. Они имеют высокое отношение поверхности к объему. Плазматическая мембрана эритроцитов имеет поверхностные белки и полисахариды, придающие группе крови. Ретикулоциты, образующиеся в мягкой внутренней части костей, называемой костным мозгом, теряют свои клеточные органеллы и попадают в кровоток в виде эритроцитов. В конце жизни эритроциты деградируют в печени и селезенке. Железо, фолиевая кислота и витамин B12 являются важными компонентами.
Железо, фолиевая кислота и витамин B12 являются важными компонентами.
Железо: гомеостатический контроль баланса железа находится в эпителии кишечника. Железо хранится в печени в виде ферритина. Железо, высвобождаемое из деградировавших эритроцитов, переносится в костный мозг с помощью белка трансферрина плазмы и включается в новые эритроциты.
Регуляция образования эритроцитов: образование эритроцитов стимулируется гормоном, называемым эритропоэтином, который секретируется в основном почками.
Анемия : Снижение способности крови переносить кислород может быть вызвано:
- Уменьшением общего числа эритроцитов
- Снижением концентрации гемоглобина на эритроцит
- Комбинацией обоих
- Серповидноклеточная анемия приводит к аномальной форме гемоглобина из-за генетической мутации. Полицитемия — избыток эритроцитов, приводящий к уменьшению кровотока в капиллярах.

(2) Лейкоциты
Различают 3 типа полиморфноядерных гранулоцитов (характеризуются наличием многодольчатых ядер и гранул): (1) эозинофилы , (2) базофилы , (4) нейтрофилы 9044 (4) нейтрофилы и 4 много), моноциты и лимфоциты. Все лейкоциты образуются в костном мозге. Эти клетки участвуют в иммунной защите.
ТромбоцитыТромбоциты представляют собой бесцветные фрагменты клеток, которые попадают в кровоток при отщеплении цитоплазматических частей клеток костного мозга, называемых мегакариоцитами.Их основная функция заключается в процессе свертывания крови.
Регуляция образования клеток крови
У детей костный мозг большинства костей вырабатывает клетки крови, тогда как у взрослых клетки крови вырабатываются только в костях верхней части тела. Все клетки крови происходят из одной популяции клеток костного мозга, называемых плюрипотентными гемопоэтическими стволовыми клетками. Эти клетки могут делиться на (1) плюрипотентные стволовые клетки , (2) лимфоидные стволовые клетки , дающие начало лимфоцитам, или (3) миелоидные стволовые клетки , дающие начало всем другим типам клеток крови.
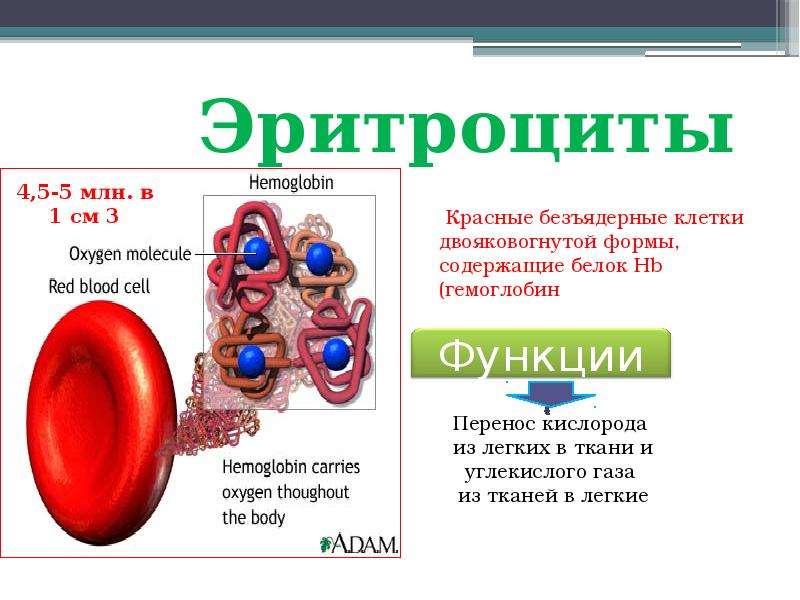 Деление и дифференцировка этих клеток регулируются белковыми гормонами и паракринными агентами, которые в совокупности называются гемопоэтическими факторами роста (HGF).
Деление и дифференцировка этих клеток регулируются белковыми гормонами и паракринными агентами, которые в совокупности называются гемопоэтическими факторами роста (HGF).Конструкция сердечно-сосудистой системы
Быстрый кровоток в одном направлении называется объемным потоком . Это производится насосной деятельностью сердца. Высокая разветвленность сосудов обеспечивает близость всех клеток к некоторым капиллярам. Питательные вещества и конечные продукты метаболизма перемещаются между капиллярной кровью и интерстициальной жидкостью путем диффузии.
Сердце продольно разделено на 2 половины: левую и правую, и каждая половина содержит две камеры: верхнее предсердие и нижний желудочек. Предсердие на каждой стороне соединяется с желудочком на этой стороне, но между двумя предсердиями или двумя желудочками нет связи. Кровь откачивается из сердца по одному набору сосудов и возвращается к сердцу по другому. Сосуды, несущие кровь от сердца, называются артериями , а сосуды, несущие кровь к сердцу, называются венами .

Легочное кровообращение (через легкие для оксигенации) Системное кровообращение (к конечностям и обратно)
Давление, поток и сопротивление
Кровь течет из области низкого давления в область высокого скорость кровотока (F) дается
F=Δp/R
где:
Δp – разница давлений между двумя точками
R – сопротивление потоку
R, в свою очередь, определяется вязкостью кровь и длина & радиус кровеносных сосудов.
В большинстве физиологических состояний изменение радиуса кровеносных сосудов регулирует кровоток. (т.е. вазоконстрикция, вазодилатация)Анатомия сердца
Сердце представляет собой мышцу, заключенную в мешок, называемый перикардом . Стенки сердца состоят из клеток сердечной мышцы, называемых миокардом. Тонкий слой клеток, называемых эндотелиальными клетками, выстилает внутреннюю поверхность. Между предсердием и желудочком с каждой стороны расположены атриовентрикулярные (АВ) клапаны.
 Правый атриовентрикулярный клапан называется трехстворчатым клапаном, а левый атриовентрикулярный клапан называется митральным клапаном. Клапан в месте открытия правого желудочка в легочную артерию называется легочным клапаном. Клапан, в котором левый желудочек входит в аорту, называется аортальным клапаном, а эти два клапана также называют полулунными клапанами. Эти клапаны позволяют крови течь только в одном направлении, а их открытие и закрытие является пассивным процессом, возникающим в результате перепада давления на клапанах.
Правый атриовентрикулярный клапан называется трехстворчатым клапаном, а левый атриовентрикулярный клапан называется митральным клапаном. Клапан в месте открытия правого желудочка в легочную артерию называется легочным клапаном. Клапан, в котором левый желудочек входит в аорту, называется аортальным клапаном, а эти два клапана также называют полулунными клапанами. Эти клапаны позволяют крови течь только в одном направлении, а их открытие и закрытие является пассивным процессом, возникающим в результате перепада давления на клапанах.Сердечная мышца
Клетки сердечной мышцы поперечнополосатые. Десмосомы и щелевые соединения в структурах, называемых вставочными дисками, соединяют соседние клетки. Некоторые клетки не участвуют в сокращении, но они образуют проводящую систему, которая инициирует сердцебиение и распространяет его по всему сердцу. Эти мышечные клетки, очевидно, жизненно важны, и они иннервируются большим количеством симпатических волокон, которые выделяют норадреналин, и парасимпатических волокон, которые выделяют ацетилхолин.
 Кровоснабжение клеток сердечной мышцы осуществляется коронарными артериями и коронарными венами соответственно. Кровь, прокачиваемая через камеры, не обменивается веществами с клетками сердечной мышцы.
Кровоснабжение клеток сердечной мышцы осуществляется коронарными артериями и коронарными венами соответственно. Кровь, прокачиваемая через камеры, не обменивается веществами с клетками сердечной мышцы.Координация сердцебиения
Последовательность возбуждения. Группа нервных клеток, называемая синоатриальным (СА) узлом в правом предсердии, деполяризуется первой. Скорость разряда узла SA определяет частоту сердечных сокращений. Деполяризация быстро распространяется на левое предсердие, и оба предсердия сокращаются одновременно.Затем потенциал действия распространяется на желудочки после небольшой задержки через атриовентрикулярный (АВ) узел, расположенный в основании правого предсердия. Задержка потенциала действия позволяет завершить сокращение предсердий до сокращения желудочков. Затем потенциал распространяется на желудочки через пучок Гиса (атриовентрикулярный пучок) и волокна Пуркинье, и оба желудочка сокращаются одновременно. Способность СА-узла к спонтанному ритмичному самовозбуждению является результатом постепенной деполяризации (пейсмекерного потенциала) клеток.
 Деполяризация, в свою очередь, происходит, когда Na+-каналы вновь открываются во время фазы реполяризации предыдущего потенциала.
Деполяризация, в свою очередь, происходит, когда Na+-каналы вновь открываются во время фазы реполяризации предыдущего потенциала.Электрокардиограмма. Электрические события в сердце могут быть косвенно зарегистрированы на поверхности кожи по токам, генерируемым во внеклеточной жидкости. Запись ЭКГ (ЭКГ) должна состоять из 3-х отклонений: (1) зубец Р – деполяризация предсердий, (2) комплекс QRS – деполяризация желудочков и (3) зубец Т – реполяризация желудочков.Длительный рефрактерный период клеток сердечной мышцы ограничивает повторное возбуждение клеток сердечного нерва, подавляя тем самым столбняк.
Механические события сердечного цикла
Сердечный цикл делится на две фазы:
- Систола: фаза сокращения желудочков и выброса крови. Во время первой части фазы систолы желудочки сокращаются, когда все клапаны еще закрыты, и поэтому кровь не выбрасывается. Этот период называется изоволюметрическим сокращением желудочков.Объем крови, выбрасываемый из каждого желудочка, называется ударным объемом (УО).
 Количество крови, остающееся после выброса, называется конечно-систолическим объемом (КСО).
Количество крови, остающееся после выброса, называется конечно-систолическим объемом (КСО). - Диастола : фаза, когда желудочки расслабляются и кровь наполняет камеры. Во время первой части диастолы желудочки расслабляются, а все клапаны еще закрыты. Этот период называется изоволюметрической релаксацией желудочков. Количество крови в желудочке в конце диастолы называется конечно-диастолическим объемом (КДО).
SV = EDV – ESV
Сердечный выброс
Чтобы найти объем крови, перекачиваемый каждым желудочком в минуту:
CO = HR x SV Сердечный выброс равен частоте сердечных сокращений, умноженной на ударный объем.Контроль сердечного ритма. Узел SA иннервируется вегетативной нервной системой. Активность парасимпатических нервов высвобождает Ач, , которые закрывают Na+-каналы и уменьшают наклон пейсмекерного потенциала, вызывая снижение частоты сердечных сокращений.Активность симпатических нервов высвобождает норадреналин, который затем открывает каналы Na+ и увеличивает наклон пейсмекерного потенциала, вызывая увеличение частоты сердечных сокращений.

Контроль ударного объема. Более сильное сокращение желудочка может привести к большему опорожнению желудочка, что приведет к увеличению ударного объема.
EDV & SV: Механизм Франка-Старлинга. Чем выше КДО, тем больше растягиваются мышцы желудочков. Это также означает более сильное сокращение.Сердечная мышца в норме не имеет оптимальной длины (lo). Таким образом, дополнительное растяжение увеличивает силу сокращения. В целом, по мере уменьшения конечного систолического объема общий ударный объем увеличивается.
Симпатические нервы высвобождают норадреналин, который может повышать сократительную способность миокарда за счет усиления инфузии кальция.
Сосудистая система
Артерии
Артерии представляют собой крупные эластичные трубки, выстланные изнутри эндотелиальными клетками. Стенки артерий состоят из соединительной ткани и гладкой мускулатуры.Во время систолы при сокращении желудочков кровь выбрасывается в артерии, растягивая стенки артерий.
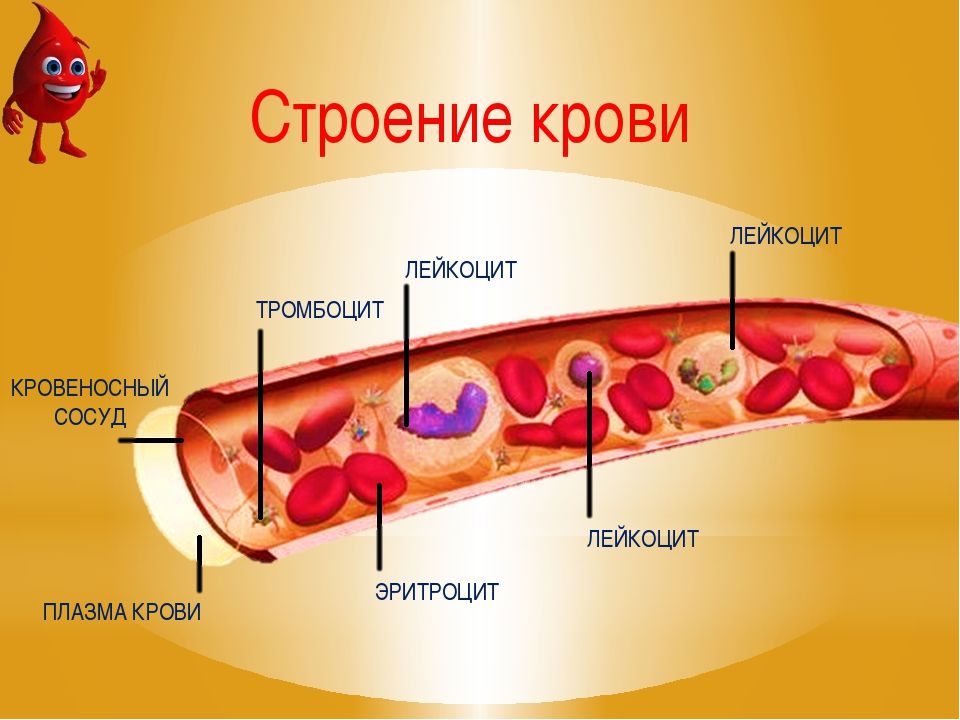 Во время диастолы стенки пассивно сокращаются, и кровь выталкивается наружу. В артериях всегда есть немного крови, чтобы держать их наполовину надутыми. Максимальное артериальное давление, достигаемое во время систолы, называется систолическим давлением (СД), а минимальное артериальное давление, достигаемое во время диастолы, называется диастолическим давлением (ДПИ). Разница между SP и DP называется пульсовым давлением (PP). Среднее давление, нагнетающее кровь в ткани, называется средним артериальным давлением (САД).
Во время диастолы стенки пассивно сокращаются, и кровь выталкивается наружу. В артериях всегда есть немного крови, чтобы держать их наполовину надутыми. Максимальное артериальное давление, достигаемое во время систолы, называется систолическим давлением (СД), а минимальное артериальное давление, достигаемое во время диастолы, называется диастолическим давлением (ДПИ). Разница между SP и DP называется пульсовым давлением (PP). Среднее давление, нагнетающее кровь в ткани, называется средним артериальным давлением (САД).Артериолы
Артериолы содержат гладкие мышцы, которые могут расслабляться для увеличения радиуса сосуда (вазодилатация) или сокращаться для уменьшения радиуса сосуда (вазоконстрикция). Он регулирует кровоток через орган. Это можно рассчитать по следующей формуле:
Forgan = MAP / ResistanceorganМестный контроль. Местный контроль — это механизмы, независимые от гормонов и нервов. Гиперемия возникает, когда кровоток в органе увеличивается за счет расширения артериол в ответ на повышение метаболической активности, что вызывает локальные изменения, такие как снижение О 2 или повышение СО 2 и Н + .
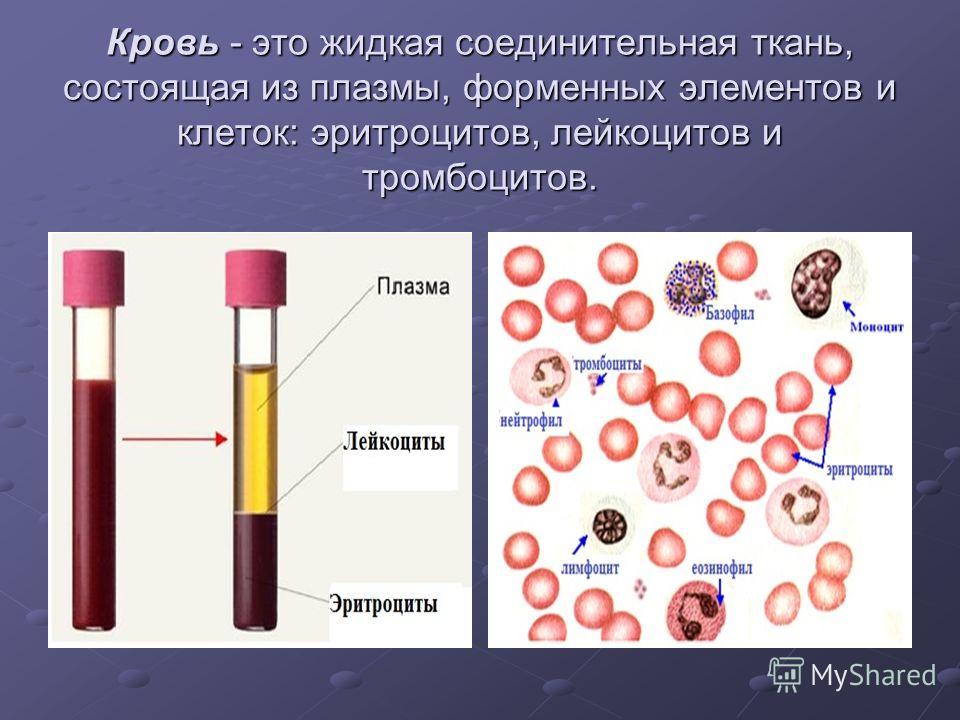
Внешний контроль. Симпатические нервы обеспечивают богатую подачу импульсов к артериолам. При активации артериолы выделяют норадреналин, вызывая вазоконстрикцию. Гормоны, такие как вазопрессин (из задней доли гипофиза) и ангиотензин II (из печени), сужают артериолы.
Капилляры
Капилляры пронизывают каждую ткань организма и обеспечивают прямой доступ к клеткам для обмена питательными веществами и конечными продуктами метаболизма.
Анатомия капиллярной сети. Капилляр представляет собой тонкостенную трубку эндотелиальных клеток толщиной в один слой, покоящуюся на базальной мембране без какой-либо окружающей мышечной или эластичной ткани. Эндотелиальные клетки отделены друг от друга узкими, заполненными водой пространствами, называемыми межклеточными щелями.
Скорость капиллярного кровотока. Скорость кровотока уменьшается, когда кровь проходит через огромную площадь поперечного сечения капилляра.

Диффузия и обмен через стенку капилляра. Существует три основных механизма, с помощью которых вещества перемещаются через стенки капилляров для входа или выхода из интерстициальной жидкости:
- Диффузия является единственным важным средством, с помощью которого может происходить чистое движение питательных веществ, кислорода и конечных продуктов метаболизма.Межклеточные щели позволяют проходить полярным молекулам. Капилляры головного мозга, однако, плотные, без межклеточных щелей. Капилляры печени неплотные с большими расщелинами для движения веществ. Градиент транскапиллярной диффузии создается за счет использования или производства вещества.
- Транспорт везикул обеспечивает прохождение молекул посредством эндо- и экзоцитоза.
- Объемный поток позволяет безбелковой плазме перемещаться из капилляров в интерстициальную жидкость благодаря гидростатическому давлению.Этому противостоит осмотическая сила, возникающая из-за различий в концентрации белка, которая имеет тенденцию перемещать интерстициальную жидкость в капилляры.
 Объемный поток также служит для распределения внеклеточной жидкости.
Объемный поток также служит для распределения внеклеточной жидкости.
Чистое давление фильтрации (NFP) можно рассчитать по формуле:
NFP = Pc – PIF – πP + πIF
Где:
Pc = капиллярное гидростатическое давление (в пользу капиллярного гидростатического давления) PIF = гидростатическое давление интерстициальной жидкости (способствует движению жидкости в капилляры)
πP = осмотическая сила, обусловленная концентрацией белков плазмы (благоприятствующая движению жидкости в капилляры)
πIF = осмотическая сила, обусловленная концентрацией белка интерстициальной жидкости (благоприятствующая движению жидкости из капилляров) )Эти четыре фактора называются силами Старлинга.
Вены
Вены представляют собой тонкостенные сосуды с низким сопротивлением, которые несут кровь от тканей к сердцу.
Детерминанты венозного давления. Общий объем крови является важным фактором, определяющим венозное давление.
 В любой момент времени большая часть крови находится в венах. Стенки вен более эластичны и поэтому могут вмещать большие объемы крови при относительно небольшом повышении давления. Стенки содержат гладкие мышцы, иннервируемые симпатическими нейронами, которые выделяют норадреналин.Это сужает сосуды, которые повышают давление и гонят больше крови. При сокращении скелетной мышцы вены в мышце сдавливаются. Это уменьшает диаметр вен, что приводит к повышению давления. Это называется накачкой скелетных мышц. Когда диафрагма опускается во время вдоха, возникает повышение давления во внутрибрюшных венах и снижение давления во внутригрудных венах, что приводит к увеличению венозного давления. Это называется дыхательный насос. Венозные клапаны препятствуют обратному току крови в венах.
В любой момент времени большая часть крови находится в венах. Стенки вен более эластичны и поэтому могут вмещать большие объемы крови при относительно небольшом повышении давления. Стенки содержат гладкие мышцы, иннервируемые симпатическими нейронами, которые выделяют норадреналин.Это сужает сосуды, которые повышают давление и гонят больше крови. При сокращении скелетной мышцы вены в мышце сдавливаются. Это уменьшает диаметр вен, что приводит к повышению давления. Это называется накачкой скелетных мышц. Когда диафрагма опускается во время вдоха, возникает повышение давления во внутрибрюшных венах и снижение давления во внутригрудных венах, что приводит к увеличению венозного давления. Это называется дыхательный насос. Венозные клапаны препятствуют обратному току крови в венах.Лимфатическая система
Лимфатическая система представляет собой сеть мелких органов (лимфатических узлов) и трубочек (лимфатических сосудов), по которым течет лимфа. Лимфа представляет собой жидкость, полученную из интерстициальной жидкости.
 Лимфатические капилляры состоят из одного слоя эндотелиальных клеток, лежащих на базальной мембране. Их водные каналы проницаемы для всех компонентов интерстициальной жидкости, включая белок. Интерстициальная жидкость поступает в эти капилляры объемным током, и жидкость течет через лимфатические узлы и заканчивается в двух лимфатических протоках, которые впадают в подключичные вены в нижней части шеи.Лимфатические сосуды переносят интерстициальную жидкость обратно в сердечно-сосудистую систему и компенсируют чистую фильтрацию из кровеносных капилляров. Кроме того, лимфатическая система обеспечивает путь, по которому жир, абсорбированный в желудочно-кишечном тракте, попадает в кровь. Инфекции, вызывающие закупорку лимфатической системы, приводят к скоплению интерстициальной жидкости, что называется отеком.
Лимфатические капилляры состоят из одного слоя эндотелиальных клеток, лежащих на базальной мембране. Их водные каналы проницаемы для всех компонентов интерстициальной жидкости, включая белок. Интерстициальная жидкость поступает в эти капилляры объемным током, и жидкость течет через лимфатические узлы и заканчивается в двух лимфатических протоках, которые впадают в подключичные вены в нижней части шеи.Лимфатические сосуды переносят интерстициальную жидкость обратно в сердечно-сосудистую систему и компенсируют чистую фильтрацию из кровеносных капилляров. Кроме того, лимфатическая система обеспечивает путь, по которому жир, абсорбированный в желудочно-кишечном тракте, попадает в кровь. Инфекции, вызывающие закупорку лимфатической системы, приводят к скоплению интерстициальной жидкости, что называется отеком.Механизм движения лимфы
Движение лимфы происходит за счет ритмичных сокращений гладких мышц, выстилающих стенки лимфатических сосудов.Сокращения вызываются растяжением стенок при попадании лимфы в систему.
 Лимфатические сосуды имеют клапаны, обеспечивающие односторонний поток. Сосуды иннервируются симпатическими нейронами, а также находятся под влиянием насоса скелетных мышц и дыхательного насоса.
Лимфатические сосуды имеют клапаны, обеспечивающие односторонний поток. Сосуды иннервируются симпатическими нейронами, а также находятся под влиянием насоса скелетных мышц и дыхательного насоса.Регуляция системного артериального давления
Среднее артериальное давление определяется сердечным выбросом и общим периферическим сопротивлением (ОПСС). TPR представляет собой сумму сопротивлений кровотоку, оказываемых всеми системными кровеносными сосудами.
MAP = CO x TPR
Сопротивление артериол является основным фактором, определяющим TPR. Любое отклонение в MAP вызывает гомеостатические рефлексы, так что CO или TPR изменяются, чтобы минимизировать отклонение.
Барорецепторные рефлексы
- Артериальные барорецепторы. Это краткосрочные регуляторы MAP. Рецепторы давления присутствуют в каротидном синусе на шее, дуге аорты (барорецепторы дуги аорты), легочных сосудах, стенке сердца и крупных системных венах.
 Афферентные нейроны, скорость возбуждения которых положительно коррелирует с MAP, от этих рецепторов направляются к стволу мозга.
Афферентные нейроны, скорость возбуждения которых положительно коррелирует с MAP, от этих рецепторов направляются к стволу мозга. - Медуллярно-сердечно-сосудистый центр. Это первичный интегрирующий центр барорецепторных рефлексов в продолговатом мозге. Когда артериальные барорецепторы уменьшают свою разрядку в результате меньшего среднего артериального давления, симпатический отток увеличивается, увеличивая частоту сердечных сокращений, сократимость желудочков и вазоконстрикцию. Он также вызывает повышенную секрецию ангиотензина II и вазопрессина, которые сужают артериолы.
Объем крови
Длительное регулирование среднего артериального давления зависит от объема крови. Увеличение MAP уменьшает объем крови за счет увеличения экскреции соли и воды почками, что приводит к снижению MAP.
Кровоизлияние и другие причины гипотонии
Гипотензия — это низкое кровяное давление из-за низкого объема крови. УО, СВ, САД снижаются как прямой результат кровоизлияния, и артериальные барорецепторные рефлексы работают на их восстановление до нормы.
 ЧСС и TPR увеличиваются как рефлекторные ответы из-за увеличения симпатического оттока.Интерстициальная жидкость перемещается в сосудистую систему из-за снижения капиллярного давления. В долгосрочной перспективе изменяются потребление жидкости и экскреция почками, а также стимулируется эритропоэз для возмещения объема крови. Потеря большого количества бесклеточной внеклеточной жидкости из-за потоотделения, рвоты, диареи и т. д. также вызывает аналогичные симптомы и реакции. Гипотония может вызвать обморок. Гипотензия может быть показателем недостаточности вегетативной нервной системы.
ЧСС и TPR увеличиваются как рефлекторные ответы из-за увеличения симпатического оттока.Интерстициальная жидкость перемещается в сосудистую систему из-за снижения капиллярного давления. В долгосрочной перспективе изменяются потребление жидкости и экскреция почками, а также стимулируется эритропоэз для возмещения объема крови. Потеря большого количества бесклеточной внеклеточной жидкости из-за потоотделения, рвоты, диареи и т. д. также вызывает аналогичные симптомы и реакции. Гипотония может вызвать обморок. Гипотензия может быть показателем недостаточности вегетативной нервной системы.Шок
Повреждение тканей или органов из-за снижения кровотока называется шоком.
- Гиповолемический шок, вызванный уменьшением объема крови из-за кровоизлияния или потери жидкости
- Шок с низким сопротивлением, вызванный снижением TPR из-за избыточного высвобождения сосудорасширяющих средств, как при аллергии и инфекции
- Кардиогенный шок из-за к снижению СО (сердечного выброса), как при сердечном приступе.

Уменьшение эффективного объема циркулирующей крови при переходе из горизонтального положения в вертикальное.В горизонтальном положении все кровеносные сосуды находятся на одном уровне и почти все давление обусловлено сердечным выбросом. В вертикальном положении в каждой точке возникает дополнительное давление, равное весу столба крови от сердца до этой точки. Это приводит к растяжению кровеносных сосудов из-за скопления крови и усилению капиллярной фильтрации в нижних частях тела. Действие гравитации может быть компенсировано сокращением скелетных мышц ног.
Упражнения
По мере увеличения CO увеличивается приток крови к мышцам и коже (для рассеивания тепла).СВ повышается за счет значительного увеличения ЧСС, вызванного усилением активности в СА-узле, и небольшого увеличения УО, вызванного повышенной сократимостью желудочков, опосредованной симпатической активностью. Также наблюдается увеличение КДО и вступает в действие механизм Франка-Старлинга.
 Венозному возврату способствуют: (1) повышенная активность насоса скелетных мышц и (2) повышенная активность дыхательного насосного вдоха (из-за увеличения глубины и частоты вдоха). Механизмы контроля этих сердечно-сосудистых изменений включают прямую регуляцию, активную гиперемию, сброс артериальных барорецепторов.
Венозному возврату способствуют: (1) повышенная активность насоса скелетных мышц и (2) повышенная активность дыхательного насосного вдоха (из-за увеличения глубины и частоты вдоха). Механизмы контроля этих сердечно-сосудистых изменений включают прямую регуляцию, активную гиперемию, сброс артериальных барорецепторов.Максимальное потребление кислорода и тренировка
Потребление кислорода увеличивается пропорционально величине упражнений до точки максимального потребления кислорода (VO2max). После достижения VO2max любое дальнейшее увеличение работы может лишь кратковременно поддерживаться анаэробным метаболизмом. VO2max ограничивается: (1) CO, (2) способностью дыхательной системы доставлять кислород в кровь и (3) способностью мышц использовать кислород. В норме VO2max определяется сердечным выбросом.
Гипертония
Повышенное артериальное давление обычно связано с увеличением TPR в результате уменьшения радиуса артериол. Почечная гипертензия возникает в результате повышенной секреции ренина, который вырабатывает ангиотензин II – вазоконстриктор.
 Гипертония приводит к увеличению мышечной массы левого желудочка (гипертрофия левого желудочка), так как ему приходится работать против повышенного артериального давления. Это может снизить сократимость, что приведет к сердечной недостаточности.
Гипертония приводит к увеличению мышечной массы левого желудочка (гипертрофия левого желудочка), так как ему приходится работать против повышенного артериального давления. Это может снизить сократимость, что приведет к сердечной недостаточности.Сердечная недостаточность
При сердечной недостаточности сердце не может перекачивать достаточное количество CO.При диастолической дисфункции стенка желудочка имеет сниженную податливость и сниженную способность адекватно наполняться, что приводит к снижению КДО и, следовательно, к уменьшению УО. Систолическая дисфункция возникает в результате повреждения миокарда и приводит к снижению сократительной способности сердца и снижению УО. Адаптивные рефлексы, противодействующие снижению СО, приводят к (1) задержке жидкости и могут вызвать отек — один в легком может ухудшить газообмен, и (2) повышенный TPR затрудняет работу сердца.
Ишемическая болезнь сердца и сердечные приступы
При ишемической болезни сердца изменения в коронарных артериях вызывают недостаточный приток крови (ишемию) к сердцу, что приводит к повреждению миокарда (инфаркт миокарда или сердечный приступ).
 Боли в груди, связанные с этим, называются стенокардией. Фибрилляция желудочков вызывает аномальную проводимость импульса поврежденными клетками миокарда, что приводит к нескоординированным сокращениям желудочков. Основной причиной ишемической болезни сердца является атеросклероз — утолщение стенки артерии из-за (1) аномальных гладких мышц, (2) отложений холестерина или (3) плотных слоев соединительной ткани. Утолщенная стенка уменьшает кровоток, а также высвобождает сосудосуживающие вещества. Атеросклероз церебральной артерии может привести к локальному повреждению головного мозга – инсульту или обратимому неврологическому дефициту, называемому транзиторной ишемической атакой (ТИА).Коронарный тромбоз – это полная окклюзия кровеносного сосуда тромбом.
Боли в груди, связанные с этим, называются стенокардией. Фибрилляция желудочков вызывает аномальную проводимость импульса поврежденными клетками миокарда, что приводит к нескоординированным сокращениям желудочков. Основной причиной ишемической болезни сердца является атеросклероз — утолщение стенки артерии из-за (1) аномальных гладких мышц, (2) отложений холестерина или (3) плотных слоев соединительной ткани. Утолщенная стенка уменьшает кровоток, а также высвобождает сосудосуживающие вещества. Атеросклероз церебральной артерии может привести к локальному повреждению головного мозга – инсульту или обратимому неврологическому дефициту, называемому транзиторной ишемической атакой (ТИА).Коронарный тромбоз – это полная окклюзия кровеносного сосуда тромбом.Гемостаз – предотвращение кровопотери
Гемостаз – это остановка кровотечения из мелких сосудов. Венозное кровотечение приводит к менее быстрой кровопотере, потому что в венах более низкое кровяное давление. Скопление крови в тканях в результате кровотечения называется гематомой.
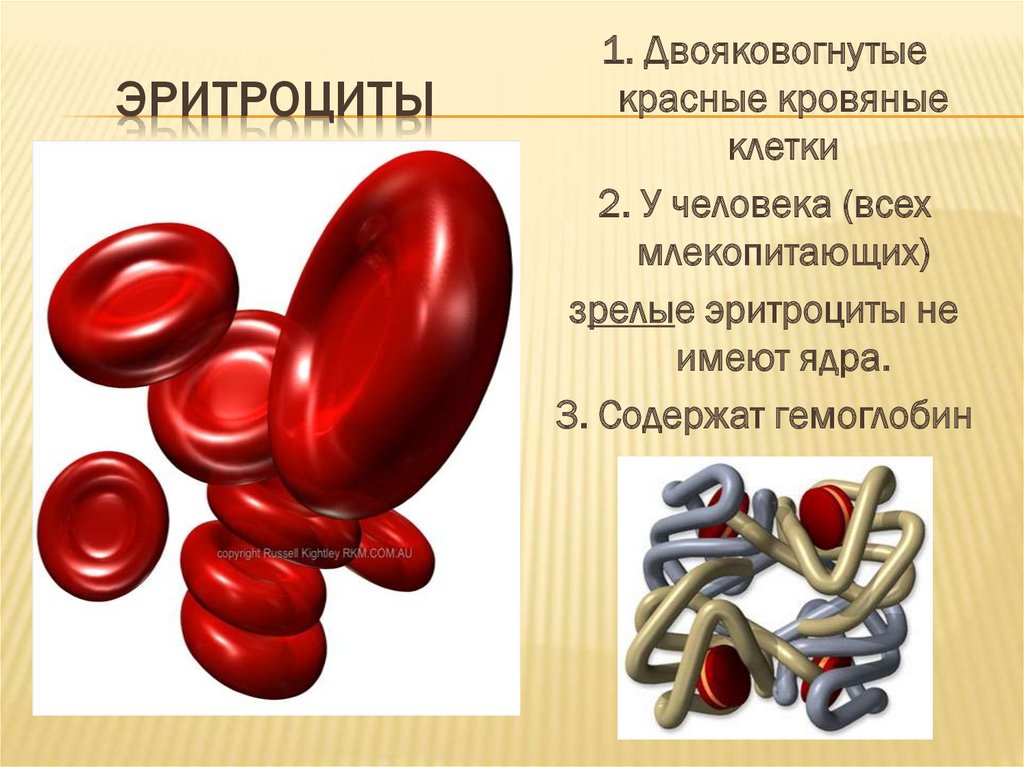 Когда кровеносный сосуд разрывается, он сужается, и противоположные эндотелиальные поверхности сосуда слипаются, замедляя отток.За ним следуют другие процессы, включая свертывание крови.
Когда кровеносный сосуд разрывается, он сужается, и противоположные эндотелиальные поверхности сосуда слипаются, замедляя отток.За ним следуют другие процессы, включая свертывание крови.Формирование тромбоцитарной пробки
Повреждение сосуда обнажает подлежащий коллаген соединительной ткани, а тромбоциты связываются с коллагеном через посредника, называемого фактором фон Виллебранда (vWF) – белком плазмы, секретируемым эндотелиальными клетками и тромбоцитами. Связывание тромбоцитов с коллагеном вызывает высвобождение секрета тромбоцитов, который изменяет форму и поверхностные белки тромбоцитов (активация тромбоцитов), заставляя их слипаться (агрегация тромбоцитов) и создавая тромбоцитарную пробку.Тромбоцитарная пробка действует как первичный герметик. Пробка не расширяется от поврежденного эндотелия, потому что интактный эндотелий синтезирует и высвобождает простациклин (простагландин 12, PGI2), который ингибирует агрегацию тромбоцитов.
Свертывание крови: образование сгустка
Кровь превращается в твердый гель, называемый сгустком или тромбом, который состоит в основном из белка фибрина.
 Он поддерживает и укрепляет пробку тромбоцитов. Белок плазмы протромбин превращается в фермент тромбин, который затем катализирует образование фибрина из фибриногена.Тромбоциты необходимы для образования сгустка, поскольку они обеспечивают поверхность, на которой происходят многие реакции. Витамин К необходим в качестве предшественника для производства протромбина и других факторов свертывания крови. Плазменный кальций также необходим для этого процесса.
Он поддерживает и укрепляет пробку тромбоцитов. Белок плазмы протромбин превращается в фермент тромбин, который затем катализирует образование фибрина из фибриногена.Тромбоциты необходимы для образования сгустка, поскольку они обеспечивают поверхность, на которой происходят многие реакции. Витамин К необходим в качестве предшественника для производства протромбина и других факторов свертывания крови. Плазменный кальций также необходим для этого процесса.Противосвертывающие системы
Фибринолитическая (тромболитическая) система удаляет тромб после восстановления сосуда. Активаторы плазминогена активируют профермент плазминогена в фермент плазмин, который расщепляет фибрин для растворения сгустка.
Распространенные препараты против свертывания крови
Аспирин, гепарин, стрептокиназа
СледующийМолекулярная биология гематоэнцефалического и гемато-ликворного барьеров: сходства и различия | Жидкости и барьеры ЦНС
- Систола: фаза сокращения желудочков и выброса крови. Во время первой части фазы систолы желудочки сокращаются, когда все клапаны еще закрыты, и поэтому кровь не выбрасывается. Этот период называется изоволюметрическим сокращением желудочков.Объем крови, выбрасываемый из каждого желудочка, называется ударным объемом (УО).
- 1.

Кроун С., Кристенсен О.: Электрическое сопротивление эндотелия капилляров. J Gen Physiol. 1981, 77: 349-371. 10.1085/jgp.77.4.349.
КАС пабмед Статья Google Scholar
- 2.
Брэдбери М.В.: Концепция гематоэнцефалического барьера. 1979, Чичестер: Wiley
Google Scholar
- 3.
Begley DJ, Brightman MW: Структурные и функциональные аспекты гематоэнцефалического барьера. Прог Наркотик Рес. 2003, 61: 39-78.
КАС пабмед Google Scholar
- 4.
Кущинский В., Полсон О.Б.: Капиллярное кровообращение в головном мозге. Cerebrovasc Brain Metab Rev. 1992, 4: 261-286.
КАС пабмед Google Scholar
- 5.
Del Bigio MR: Эпендима: защитный барьер между мозгом и спинномозговой жидкостью. Глия. 1995, 14: 1-13. 10.1002/глиа.440140102.
КАС пабмед Статья Google Scholar
- 6.

Saito Y, Wright EM: Регуляция транспорта бикарбоната через мембрану щеточной каймы хориоидального сплетения лягушки-быка.Дж. Физиол. 1984, 350: 327-342.
Центральный пабмед КАС пабмед Статья Google Scholar
- 7.
Welch K, Araki H: Особенности сосудистого сплетения кошки, изученные in vitro. Жидкая среда головного мозга. Под редакцией: Cserr HF, Fenstermacher JD, Fencl V. 1975, New York: Academic Press, 157–165.
Google Scholar
- 8.
Скотт Д.Е., Козловский Г.П., Шеридан М.Н.: Сканирующая электронная микроскопия в ультраструктурном анализе церебрально-желудочковой системы млекопитающих.Int Rev Cytol. 1974, 37: 349-388. полный текст.
КАС пабмед Статья Google Scholar
- 9.
Keep RF, Jones HC: Морфометрическое исследование развития сосудистого сплетения бокового желудочка, капилляров сосудистого сплетения и эпендимы желудочка у крыс.
-С. Dev Brain Res. 1990, 56: 47-53. 10.1016/0165-3806(90)
Dev Brain Res. 1990, 56: 47-53. 10.1016/0165-3806(90)КАС Статья Google Scholar
- 10.
Хокинс Б.Т., Дэвис Т.П.: Нервно-сосудистая единица гематоэнцефалического барьера в норме и при патологии. Pharmacol Rev. 2005, 57: 173-185. 10.1124/пр.57.2.4.
КАС пабмед Статья Google Scholar
- 11.
Martin-Padura I, Lostaglio S, Schneemann M, Williams L, Romano M, Fruscella P, Panzeri C, Stoppacciaro A, Ruco L, Villa A, Simmons D, Dejana E: Молекула соединительной адгезии, роман член суперсемейства иммуноглобулинов, который распределяется в межклеточных соединениях и модулирует трансмиграцию моноцитов.Джей Селл Биол. 1998, 142: 117-127. 10.1083/jcb.142.1.117.
Центральный пабмед КАС пабмед Статья Google Scholar
- 12.
Gonzalez-Mariscal L, Betanzos A, Nava P, Jaramillo BE: Плотные соединительные белки.
 Прог Биофиз Мол Биол. 2003, 81: 1-44. 10.1016/С0079-6107(02)00037-8.
Прог Биофиз Мол Биол. 2003, 81: 1-44. 10.1016/С0079-6107(02)00037-8.КАС пабмед Статья Google Scholar
- 13.
Li Y, Fanning AS, Anderson JM, Lavie A: Структура консервативного цитоплазматического С-концевого домена окклюдина: идентификация поверхности связывания ZO-1.Дж Мол Биол. 2005, 352: 151-164. 10.1016/j.jmb.2005.07.017.
КАС пабмед Статья Google Scholar
- 14.
Фельдман Г.Дж., Маллин Дж.М., Райан М.П.: Окклюдин: структура, функция и регулирование. Adv Drug Deliv Rev. 2005, 57: 883-917. 10.1016/j.addr.2005.01.009.
КАС пабмед Статья Google Scholar
- 15.
Андреева А.Ю., Пионтек Дж., Блазиг И.Е., Утепбергенов Д.И.: Сборка плотных контактов регулируется антагонизмом традиционных и новых изоформ протеинкиназы С.Int J Biochem Cell Biol. 2006, 38: 222-233.
КАС пабмед Google Scholar
- 16.

Сайтоу М., Фурусе М., Сасаки Х., Шульцке Д., Фромм М., Такано Х., Нода Т., Цукита С.: Сложный фенотип мышей, лишенный окклюдина, компонента нитей плотного соединения. Мол Биол Селл. 2000, 11: 4131-4142.
Центральный пабмед КАС пабмед Статья Google Scholar
- 17.
Сайтоу М., Фудзимото К., Дои Ю., Ито М., Фуджимото Т., Фурусэ М., Такано Х., Нода Т., Цукита С. Эмбриональные стволовые клетки с дефицитом окклюдина могут дифференцироваться в поляризованные эпителиальные клетки, несущие плотные контакты. Джей Селл Биол. 1998, 141: 397-408. 10.1083/jcb.141.2.397.
Центральный пабмед КАС пабмед Статья Google Scholar
- 18.
Bamforth SD, Kniesel U, Wolburg H, Engelhardt B, Risau W: Доминантный мутант окклюдина нарушает структуру и функцию плотных контактов.Дж. Клеточные науки. 1999, 112: 1879-1888.
КАС пабмед Google Scholar
- 19.

Terai T, Nishimura N, Kanda I, Yasui N, Sasaki T: JRAB/MICAL-L2 представляет собой соединительный Rab13-связывающий белок, опосредующий эндоцитарную рециркуляцию окклюдина. Мол Биол Селл. 2006, 17: 2465-2475. 10.1091/mbc.E05-09-0826.
Центральный пабмед КАС пабмед Статья Google Scholar
- 20.
Romanitan MO, Popescu BO, Winblad B, Bajenaru OA, Bogdanovic N: Окклюдин сверхэкспрессируется при болезни Альцгеймера и сосудистой деменции. J Cell Mol Med. 2007, 11: 569-579. 10.1111/j.1582-4934.2007.00047.x.
Центральный пабмед КАС пабмед Статья Google Scholar
- 21.
Heiskala M, Peterson PA, Yang Y: Роль белков суперсемейства клаудина в парацеллюлярном транспорте. Движение. 2001, 2: 93-98. 10.1034/j.1600-0854.2001.020203.х.
КАС пабмед Статья Google Scholar
- 22.
Piontek J, Winkler L, Wolburg H, Muller SL, Zuleger N, Piehl C, Wiesner B, Krause G, Blasig IE: Формирование плотного соединения: детерминанты гомофильного взаимодействия между классическими клаудинами.
 FASEB J. 2008, 22: 146-158. 10.1096/fj.07-8319com.
FASEB J. 2008, 22: 146-158. 10.1096/fj.07-8319com.КАС пабмед Статья Google Scholar
- 23.
Nitta T, Hata M, Gotoh S, Seo Y, Sasaki H, Hashimoto N, Furuse M, Tsukita S: Селективное по размеру ослабление гематоэнцефалического барьера у мышей с дефицитом клаудина-5. Джей Селл Биол. 2003, 161: 653-660. 10.1083/jcb.200302070.
Центральный пабмед КАС пабмед Статья Google Scholar
- 24.
Ruffer C, Gerke V: С-концевой цитоплазматический хвост клаудинов 1 и 5, но не его PDZ-связывающий мотив, необходим для апикальной локализации в эпителиальных и эндотелиальных плотных соединениях.Eur J Cell Biol. 2004, 83: 135-144. 10.1078/0171-9335-00366.
ПабМед Статья Google Scholar
- 25.
Маттер К., Балда М.С.: Дырчатый барьер: клаудины и регуляция проницаемости эндотелия головного мозга.
 Джей Селл Биол. 2003, 161: 459-460. 10.1083/jcb.200304039.
Джей Селл Биол. 2003, 161: 459-460. 10.1083/jcb.200304039.Центральный пабмед КАС пабмед Статья Google Scholar
- 26.
Van Itallie CM, Anderson JM: Роль клаудинов в определении селективности парацеллюлярного заряда.Proc Am Thorac Soc. 2004, 1: 38-41. 10.1513/пат.2306013.
КАС пабмед Статья Google Scholar
- 27.
Wolburg H, Wolburg-Buchholz K, Liebner S, Engelhardt B: Клаудин-1, клаудин-2 и клаудин-11 присутствуют в плотных соединениях эпителия сосудистого сплетения мыши. Нейроски Летт. 2001, 307: 77-80. 10.1016/С0304-3940(01)01927-9.
КАС пабмед Статья Google Scholar
- 28.
Wolburg H, Noell S, Wolburg-Buchholz K, Mack AF, Fallier-Becker P: Агрин, аквапорин-4 и полярность астроцитов как важная особенность гематоэнцефалического барьера. Нейробиолог. 2009, 15: 180-193.
 10.1177/1073858408329509.
10.1177/1073858408329509.КАС пабмед Статья Google Scholar
- 29.
Goldmann EE: Vitalfarbung am Zentralnervensystem. Abh preuss Akad Wiss Phys-Math. 1913, 1: 1-60.
Google Scholar
- 30.
Wolburg H, Paulus W: Сосудистое сплетение: биология и патология. Акта Нейропатол. 2010, 119: 75-88. 10.1007/s00401-009-0627-8.
ПабМед Статья Google Scholar
- 31.
Sandri C, Akert K, Bennett MV: Соединительные комплексы и вариации щелевых соединений между эпендимальными клетками спинного мозга костистого Sternarchus albifrons (Gymnotoidei). Мозг Res. 1978, 143: 27-41. 10.1016/0006-8993(78)
-3.
КАС пабмед Статья Google Scholar
Zamora AJ, Thiesson D: Плотные соединения в эпендиме спинного мозга уроделе Pleurodeles waltlii. Анат Эмбриол. 1980, 160: 263-274. 10.1007/BF00305107.
Анат Эмбриол. 1980, 160: 263-274. 10.1007/BF00305107.
КАС пабмед Статья Google Scholar
Mollgard K, Balslev Y, Lauritzen B, Saunders NR: Клеточные соединения и специализации мембран в вентрикулярной зоне (зародышевый матрикс) развивающегося мозга овцы: CSF-мозговой барьер. J Нейроцитол.1987, 16: 433-444. 10.1007/BF01668498.
КАС пабмед Статья Google Scholar
Bazzoni G, Dejana E: Эндотелиальные межклеточные соединения: молекулярная организация и роль в сосудистом гомеостазе. Physiol Rev. 2004, 84: 869-901. 10.1152/физрев.00035.2003.
КАС пабмед Статья Google Scholar
Bazzoni G, Martinez-Estrada OM, Mueller F, Nelboeck P, Schmid G, Bartfai T, Dejana E, Brockhaus M: гомофильное взаимодействие соединительной молекулы адгезии.Дж. Биол. Хим. 2000, 275: 30970-30976. 10.1074/jbc.M003946200.
10.1074/jbc.M003946200.
КАС пабмед Статья Google Scholar
Itoh M, Sasaki H, Furuse M, Ozaki H, Kita T, Tsukita S: Молекула соединительной адгезии (JAM) связывается с PAR-3: возможный механизм привлечения PAR-3 к плотным контактам. Джей Селл Биол. 2001, 154: 491-497. 10.1083/jcb.200103047.
Центральный пабмед КАС пабмед Статья Google Scholar
Suzuki A, Ishiyama C, Hashiba K, Shimizu M, Ebnet K, Ohno S: активность киназы aPKC необходима для асимметричной дифференцировки преждевременного соединительного комплекса во время поляризации эпителиальных клеток. Дж. Клеточные науки. 2002, 115: 3565-3573. 10.1242/jcs.00032.
КАС пабмед Статья Google Scholar
Huber JD, Egleton RD, Davis TP: Молекулярная физиология и патофизиология плотных контактов гематоэнцефалического барьера.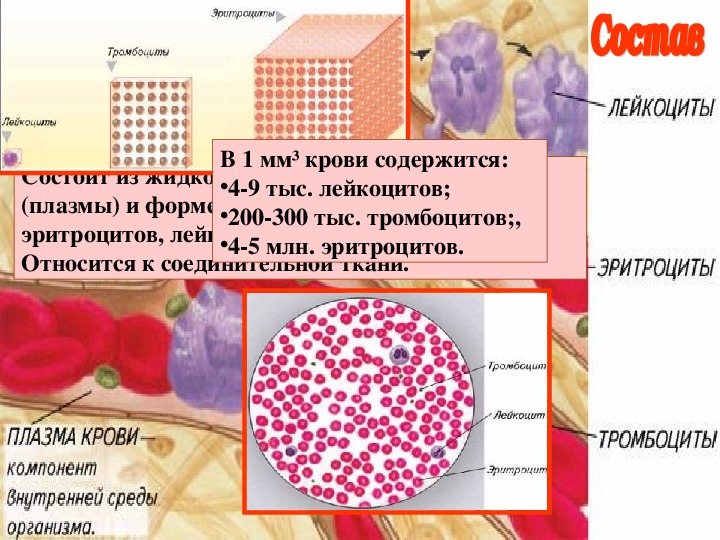 Тренды Нейроси. 2001, 24: 719-725. 10.1016/S0166-2236(00)02004-Х.
Тренды Нейроси. 2001, 24: 719-725. 10.1016/S0166-2236(00)02004-Х.
КАС пабмед Статья Google Scholar
Enerson BE, Drewes LR: Транскриптом крысиного гематоэнцефалического барьера. J Cereb Blood Flow Metab. 2006, 26: 959-973. 10.1038/sj.jcbfm.9600249.
КАС пабмед Статья Google Scholar
Koto T, Takubo K, Ishida S, Shinoda H, Inoue M, Tsubota K, Okada Y, Ikeda E: Гипоксия нарушает барьерную функцию нервных кровеносных сосудов за счет изменения экспрессии клаудина-5 в эндотелиальной ткани. клетки.Ам Джей Патол. 2007, 170: 1389-1397. 10.2353/ajpath.2007.060693.
Центральный пабмед КАС пабмед Статья Google Scholar
Kago T, Takagi N, Date I, Takenaga Y, Takagi K, Takeo S: Церебральная ишемия усиливает фосфорилирование тирозина окклюдина в капиллярах головного мозга.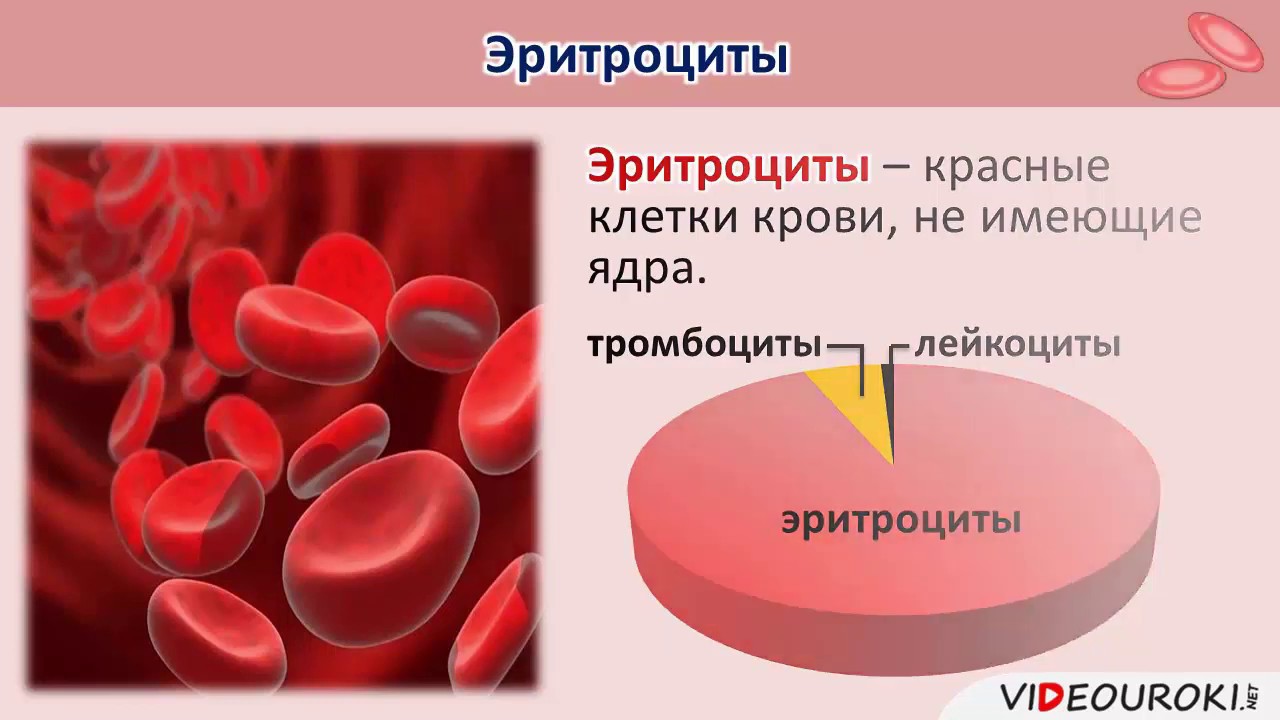 Biochem Biophys Res Commun. 2006, 339: 1197-1203. 10.1016/j.bbrc.2005.11.133.
Biochem Biophys Res Commun. 2006, 339: 1197-1203. 10.1016/j.bbrc.2005.11.133.
КАС пабмед Статья Google Scholar
Mark KS, Davis TP: Церебральные микрососудистые изменения проницаемости и плотных соединений, вызванные гипоксией-реоксигенацией. Am J Physiol Heart Circ Physiol. 2002, 282: h2485 h2494-
PubMed Central пабмед Статья Google Scholar
Fischer S, Wiesnet M, Marti HH, Renz D, Schaper W. 2004: Одновременная активация нескольких вторичных мессенджеров при индуцированной гипоксией гиперпроницаемости эндотелиальных клеток головного мозга.J Cell Physiol. 2004, 198: 359-369. 10.1002/jcp.10417.
КАС пабмед Статья Google Scholar
Рональдсон П.Т., Демарко К.М., Санчес-Коваррубиас Л., Солинский К.М., Дэвис Т.П.: трансформирующий бета-сигнальный фактор роста изменяет проницаемость субстрата и экспрессию белков плотных контактов на гематоэнцефалическом барьере во время воспалительной боли. J Cereb Blood Flow Metab. 2009, 29: 1084-98. 10.1038/jcbfm.2009.32.
J Cereb Blood Flow Metab. 2009, 29: 1084-98. 10.1038/jcbfm.2009.32.
Центральный пабмед КАС пабмед Статья Google Scholar
Lochhead JJ, McCaffrey G, Quigley CE, Finch J, DeMarco KM, Nametz N, Davis TP: Окислительный стресс увеличивает проницаемость гематоэнцефалического барьера и вызывает изменения окклюдина во время гипоксии-реоксигенации. J Cereb Blood Flow Metab. 2010, 30: 1625-1636. 10.1038/jcbfm.2010.29.
Центральный пабмед КАС пабмед Статья Google Scholar
Willis CL, Meske DS, Davis TP: Активация протеинкиназы C модулирует обратимое увеличение проницаемости кортикального гематоэнцефалического барьера и экспрессию белка плотного соединения во время гипоксии и постгипоксической реоксигенации.J Cereb Blood Flow Metab. 2010, 30: 1847-1859. 10.1038/jcbfm.2010.119.
Центральный пабмед КАС пабмед Статья Google Scholar

Lyck R, Ruderisch N, Moll AG, Steiner O, Cohen CD, Engelhardt B, Makrides V, Verrey F: Вызванные культурой изменения в транскриптоме гематоэнцефалического барьера: последствия для переносчиков аминокислот in vivo. J Cereb Blood Flow Metab. 2009, 29: 1491-1502. 10.1038/jcbfm.2009.72.
КАС пабмед Статья Google Scholar
Vestweber D: VE-кадгерин: основная молекула эндотелиальной адгезии, контролирующая клеточные соединения и формирование кровеносных сосудов. Артериосклеры Тромб Васк Биол. 2008, 28: 223-232. 10.1161/АТВБАХА.107.158014.
КАС пабмед Статья Google Scholar
Meng W, Takeichi M: Adherens junction: молекулярная архитектура и регуляция. Колд Спринг Харб Перспект Биол. 2009, 1: а002899-10.1101/cshperspect.а002899.
Центральный пабмед пабмед Статья Google Scholar

Williams MJ, Lowrie MB, Bennett JP, Firth JA, Clark P: Кадгерин-10 представляет собой новую молекулу адгезии гематоэнцефалического барьера у человека и мыши. Мозг Res. 2005, 1058: 62-72. 10.1016/j.brainres.2005.07.078.
КАС пабмед Статья Google Scholar
Lampugnani MG, Dejana E: Слипчивые соединения в эндотелиальных клетках регулируют поддержание сосудов и ангиогенез. Рез. Тромб. 2007, 120: С1-С6. 10.1016/S0049-3848(07)70124-X.
ПабМед Статья Google Scholar
Lippoldt A, Jansson A, Kniesel U, Andbjer B, Andersson A, Wolburg H, Fuxe K, Haller H: Индуцированные форболовым эфиром изменения в плотных и адгезивных соединениях в эпителии сосудистого сплетения и в эпендиме. Мозг Res. 2000, 854: 197-206. 10.1016/S0006-8993(99)02355-0.
КАС пабмед Статья Google Scholar

Дин М., Хамон И., Чимини Г.: Надсемейство переносчиков АТФ-связывающей кассеты (АВС) человека. J липидный рез. 2001, 42: 1007-1017.
КАС пабмед Google Scholar
Дуайер Д.С., Ваннуччи С.Дж., Симпсон И.А.: Экспрессия, регуляция и функциональная роль переносчиков глюкозы (GLUT) в головном мозге. Int Rev Neurobiol. 2002, 51: 159-88. 10.1016/S0074-7742(02)51005-9.
КАС пабмед Статья Google Scholar
Хокинс Р.А., О’Кейн Р.Л., Симпсон И.А., Вина Дж.Р.: Структура гематоэнцефалического барьера и его роль в транспорте аминокислот. Дж Нутр. 2006, 136: 218С-226С.
КАС пабмед Google Scholar
Kido Y, Tamai I, Okamoto M, Suzuki F, Tsuji A: Функциональное выяснение MCT1-опосредованного транспорта монокарбоновых кислот через гематоэнцефалический барьер с использованием культивируемых клеток in vitro и исследований BUI in vivo. Фарм Рез. 2000, 17: 55-62.10.1023/А:1007518525161.
Фарм Рез. 2000, 17: 55-62.10.1023/А:1007518525161.
КАС пабмед Статья Google Scholar
Spector R, Johanson CE: Транспорт витаминов и гомеостаз в мозге млекопитающих: акцент на витаминах B и E. J Neurochem. 2007, 103: 425-438. 10.1111/j.1471-4159.2007.04773.x.
КАС пабмед Статья Google Scholar
Isakovic AJ, Abbott NJ, Redzic ZB: Транспорт аденозина из мозга в кровь: исследования гематоэнцефалического барьера на крысах.Дж. Нейрохим. 2004, 90: 272-86. 10.1111/j.1471-4159.2004.02439.х.
КАС пабмед Статья Google Scholar
Redzic ZB, Biringer J, Barnes K, Baldwin SA, Al-Sarraf H, Nicola PA, Young JD, Cass CE, Barrand MA, Hladky SB: Поляризованное распределение нуклеозидных переносчиков в эндотелиальном и сосудистом сплетении головного мозга крысы эпителиальные клетки. Дж. Нейрохим. 2005, 94: 1420-1426. 10.1111/j.1471-4159.2005.03312.х.
Дж. Нейрохим. 2005, 94: 1420-1426. 10.1111/j.1471-4159.2005.03312.х.
КАС пабмед Статья Google Scholar
Исакович А.Дж., Денчич С.М., Сегал М.Б., Редзич З.Б.: Транспорт [ 14 C]гипоксантина эпителием сосудистого сплетения овец в виде монослоя в первичной культуре: Na + -зависимое и Na + -независимое поглощение апикальной мембраны и быстрого внутриклеточного метаболического превращения в нуклеотиды. Нейроски Летт. 2008, 431: 135-140. 10.1016/j.neulet.2007.11.029.
КАС пабмед Статья Google Scholar
Редзич З.Б., Малатиали С.А., Крайк Д.Д., Ракич М.Л., Исакович А.Дж.: транспорт пиримидиновых нуклеозидов и нуклеооснований через гематоэнцефалический барьер у крыс. Нейрохим Рез. 2009, 34: 566-573. 10.1007/s11064-008-9823-5.
КАС пабмед Статья Google Scholar

Симпсон И.А., Каррутерс А., Ваннуччи С.Дж.: Спрос и предложение в церебральном энергетическом обмене: роль переносчиков питательных веществ. J Cereb Blood Flow Metab. 2007, 27: 1766-1791.10.1038/sj.jcbfm.9600521.
Центральный пабмед КАС пабмед Статья Google Scholar
Tsacopoulos M, Magistretti PJ: Метаболическая связь между глией и нейронами. Дж. Нейроски. 1996, 16: 877-885.
КАС пабмед Google Scholar
Аби-Сааб В.М., Мэггс Д.Г., Джонс Т., Джейкоб Р., Шрихари В., Томпсон Дж., Керр Д., Леоне П., Кристал Дж.Х., Спенсер Д.Д., Воин М.Дж., Шервин Р.С.: Поразительные различия в глюкозе и лактате уровни между внеклеточной жидкостью головного мозга и плазмой у находящихся в сознании людей: эффекты гипергликемии и гипогликемии.J Cereb Blood Flow Metab. 2002, 22: 271-9. 10.1097/00004647-200203000-00004.
КАС пабмед Статья Google Scholar

Mueckler M, Caruso C, Baldwin SA, Panico M, Blench I, Morris HR, Allard WJ, Lienhard GE, Lodish HF: Последовательность и структура переносчика глюкозы человека. Наука. 1985, 229: 941-945. 10.1126/науки.3839598.
КАС пабмед Статья Google Scholar
Kayano T, Fukumoto H, Eddy RL, Fan YS, Byers MG, Shows TB, Bell GI: Доказательства существования семейства белков, подобных переносчикам глюкозы человека. Последовательность и локализация гена белка, экспрессируемого в скелетных мышцах плода и других тканях. Дж. Биол. Хим. 1988, 263: 15245-15248.
КАС пабмед Google Scholar
Doege H, Schürmann A, Bahrenberg G, Brauers A, Joost HG: транспортер глюкозы 8 (GLUT8): новый сахарный посредник с транспортной активностью глюкозы.Дж. Биол. Хим. 2000, 275: 16275-16280. 10.1074/jbc.275.21.16275.
КАС пабмед Статья Google Scholar

Olson AL, Pessin JE: Структура, функция и регуляция семейства генов переносчиков глюкозы у млекопитающих. Анну Рев Нутр. 1996, 16: 235-256. 10.1146/annurev.nu.16.070196.001315.
КАС пабмед Статья Google Scholar
Nishimura H, Pallardo FV, Seidner GA, Vannucci S, Simpson IA, Birnbaum MJ: Кинетика переносчиков глюкозы GLUT1 и GLUT4, экспрессируемых в ооцитах Xenopus. Дж. Биол. Хим. 1993, 268: 8514-8520.
КАС пабмед Google Scholar
Duarte JM, Morgenthaler FD, Lei H, Poitry-Yamate C, Gruetter R: Переоценка кинетики транспорта глюкозы в устойчивом состоянии мозга с помощью конформационной модели с четырьмя состояниями. Фронт Нейроэнергетика. 2009, 1: 6-
PubMed Central КАС пабмед Статья Google Scholar
Махер Ф., Ваннуччи С.Дж., Симпсон И.А.: Белки-переносчики глюкозы в головном мозге. FASEB J. 1994, 8: 1003-1010.
FASEB J. 1994, 8: 1003-1010.
КАС пабмед Google Scholar
Kumagai AK, Dwyer KJ, Pardridge WM: Дифференциальное гликозилирование переносчика глюкозы GLUT1 в капиллярах головного мозга и сосудистом сплетении. Биохим Биофиз Акта. 1994, 1193: 24-30. 10.1016/0005-2736(94)
-Х.
КАС пабмед Статья Google Scholar
Semenza GL: Кислородозависимая регуляция митохондриального дыхания фактором, индуцируемым гипоксией 1. Biochem J. 2007, 405: 1-9.
КАС пабмед Статья Google Scholar
Semenza GL: HIF-1: медиатор физиологических и патофизиологических реакций на гипоксию. J Appl Physiol. 2000, 88: 1474-1480.
КАС пабмед Google Scholar
Bruick RK, McKnight SL: консервативное семейство пролил-4-гидроксилаз, модифицирующих HIF. Наука. 2001, 294: 1337-1340. 10.1126/научн.1066373.
Наука. 2001, 294: 1337-1340. 10.1126/научн.1066373.
КАС пабмед Статья Google Scholar
Chavez JC, LaManna JC: Активация индуцируемого гипоксией фактора-1 в коре головного мозга крыс после транзиторной глобальной ишемии: потенциальная роль инсулиноподобного фактора роста-1. Дж. Нейроски. 1993, 22: 8922-8931.
Google Scholar
Вемула С., Родер К.Е., Ян Т., Бхат Г.Дж., Теккумкара Т.Дж., Аббрускато Т.Дж.: Функциональная роль натрий-зависимого транспорта глюкозы через гематоэнцефалический барьер во время кислородно-глюкозного голодания. J Pharmacol Exp Ther. 2009, 328: 487-495. 10.1124/jpet.108.146589.
Центральный пабмед КАС пабмед Статья Google Scholar
Guo X, Geng M, Du G: Переносчик глюкозы 1, распределение в головном мозге и при нервных расстройствах: его связь с транспортом нейроактивных препаратов через гематоэнцефалический барьер. Биохим Генет. 2005, 43: 175-87. 10.1007/s10528-005-1510-5.
Биохим Генет. 2005, 43: 175-87. 10.1007/s10528-005-1510-5.
КАС пабмед Статья Google Scholar
Rotstein M, Engelstad K, Yang H, Wang D, Levy B, Chung WK, De Vivo DC: Дефицит Glut1: Характер наследования определяется гаплонедостаточностью. Энн Нейрол. 2010.
Google Scholar
Клеппер Дж., Лейендекер Б.: Синдром дефицита GLUT1 — обновление 2007 года.Dev Med Child Neurol. 2007, 49: 707-16. 10.1111/j.1469-8749.2007.00707.x.
ПабМед Статья Google Scholar
Брокманн К. Расширяющийся фенотип синдрома дефицита GLUT1. Мозг Дев. 2009, 31: 545-552. 10.1016/j.braindev.2009.02.008.
ПабМед Статья Google Scholar
Wang D, Pascual JM, Yang H, Engelstad K, Mao X, Cheng J, Yoo J, Noebels JL, De Vivo DC: Мышиная модель гаплонедостаточности Glut-1.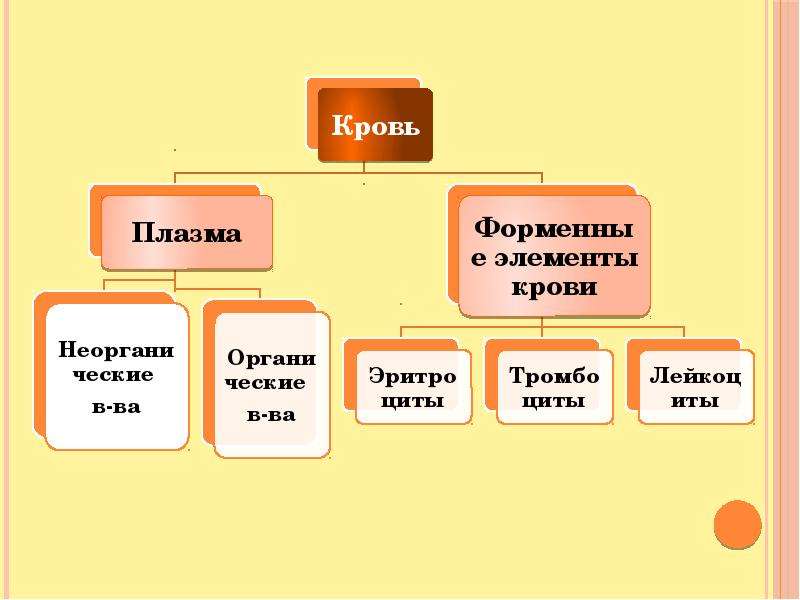 Хум Мол Жене. 1993, 15: 1169-1179. 10.1093/hmg/ddl032.
Хум Мол Жене. 1993, 15: 1169-1179. 10.1093/hmg/ddl032.
Артикул КАС Google Scholar
Mooradian AD, Chung HC, Shah GN: экспрессия GLUT-1 в головном мозге пациентов с болезнью Альцгеймера. Нейробиол Старение. 1993, 18: 469-474. 10.1016/С0197-4580(97)00111-5.
Артикул Google Scholar
Hunt A, Schonknecht P, Henze M, Seidl U, Haberkorn U, Schroder J: Снижение церебрального метаболизма глюкозы у пациентов с риском развития болезни Альцгеймера.Психиатрия рез. 155: 147-154.
Mosconi L, De Santi S, Li J, Tsui WH, Li Y, Boppana M, Laska E, Rusinek H, de Leon MJ: гипометаболизм гиппокампа предсказывает снижение когнитивных функций при нормальном старении. Нейробиол Старение. 2008, 29: 676-692. 10.1016/j.neurobiolaging.2006.12.008.
Центральный пабмед КАС пабмед Статья Google Scholar

Régina A, Morchoisne S, Borson ND, McCall AL, Drewes LR, Roux F: Факторы, высвобождаемые астроцитами, лишенными глюкозы, усиливают экспрессию и активность переносчиков глюкозы в эндотелиальных клетках головного мозга крыс.Биохим Биофиз Акта. 2001, 1540: 233-242.
ПабМед Статья Google Scholar
Yeh WL, Lin CJ, Fu WM: Повышение экспрессии транспортера глюкозы в эндотелиальных клетках головного мозга с помощью сосудистого эндотелиального фактора роста, полученного из глиомы, подвергшейся гипоксии. Мол Фармакол. 2008, 73: 170-177. 10.1124/моль.107.038851.
КАС пабмед Статья Google Scholar
Дин Р., Сегал М.Б.: Транспорт сахаров через кровоснабжаемое сосудистое сплетение овец. Дж. Физиол. 1985, 362: 245-260.
Центральный пабмед КАС пабмед Статья Google Scholar

Gerhart DZ, LeVasseur RJ, Broderius MA, Drewes LR: Локализация переносчиков глюкозы в головном мозге с использованием световой и электронной иммуноцитохимии. J Neurosci Res. 1989, 22: 464-472. 10.1002/jnr.4
413.
КАС пабмед Статья Google Scholar
Харик С.И., Калария Р.Н., Андерссон Л., Лундал П., Перри Г.: Иммуноцитохимическая локализация эритроидного переносчика глюкозы: обилие в тканях с барьерными функциями. Дж. Нейроски. 1990, 10: 3862-3872.
КАС пабмед Google Scholar
Fields HM, Rinaman L, Devaskar SU: Распределение транспортера глюкозы изоформы-3 и гексокиназы I в постнатальном мозге мышей. Мозг Res. 1999, 846: 260-264. 10.1016/S0006-8993(99)01979-4.
КАС пабмед Статья Google Scholar
Arluison M, Quignon M, Nguyen P, Thorens B, Leloup C, Penicaud L: Распределение и анатомическая локализация переносчика глюкозы 2 (GLUT2) в мозге взрослых крыс – иммуногистохимическое исследование. J Chem Neuroanat. 2004, 28: 117-36. 10.1016/j.jchemneu.2004.05.009.
J Chem Neuroanat. 2004, 28: 117-36. 10.1016/j.jchemneu.2004.05.009.
КАС пабмед Статья Google Scholar
Cornford EM, Hyman S, Cornford ME, Damian RT: переносчик глюкозы Glut1 в эндотелии сосудистого сплетения приматов. J Neuropathol Exp Neurol. 1998, 57: 404-414. 10.1097/00005072-199805000-00004.
КАС пабмед Статья Google Scholar
Pardridge WM: Мозговой метаболизм: перспектива с точки зрения гематоэнцефалического барьера. Physiol Rev. 1983, 63: 1481-1535.
КАС пабмед Google Scholar
Вуртман Р.Дж., Московиц М.А., Манро Х.Н.: Транссинаптический контроль синтеза нейронального белка. Нейронауки: Четвертая учебная программа. Под редакцией: Шмитт Ф.О., Норден Ф.Г. 1979, Кембридж: MIT Press, 897-909.
Google Scholar
Verrey F: Система L: гетеромерные обменники больших нейтральных аминокислот, участвующих в направленном транспорте. Арка Пфлюгера. 2003, 445: 529-533.
КАС пабмед Статья Google Scholar
Nakamura E, Sato M, Yang H, Miyagawa F, Harasaki M, Tomita K, Matsuoka S, Noma A, Iwai K, Minato N: тяжелая цепь 4F2 (CD98) ковалентно связана с переносчиком аминокислот и контролирует внутриклеточный перенос и мембранная топология гетеродимера 4F2. Дж. Биол. Хим. 1999, 274: 3009-3016. 10.1074/jbc.274.5.3009.
КАС пабмед Статья Google Scholar
Estévez R, Camps M, Rojas AM, Testar X, Devés R, Hediger MA, Zorzano A, Palacín M: Система транспорта аминокислот y+L/4F2hc представляет собой гетеромультимерный комплекс.FASEB J. 1998, 12: 1319-1329.
ПабМед Google Scholar
Bröer S, Bröer A, Hamprecht B: Экспрессия поверхностного антигена 4F2hc влияет на системно-L-подобную активность транспорта нейтральных аминокислот в клетках млекопитающих. Biochem J. 1997, 324: 535-541.
Biochem J. 1997, 324: 535-541.
Центральный пабмед пабмед Статья Google Scholar
Kido Y, Tamai I, Uchino H, Suzuki F, Sai Y, Tsuji A: Молекулярная и функциональная идентификация переносчиков больших нейтральных аминокислот LAT1 и LAT2 и их фармакологическая значимость для гематоэнцефалического барьера.Дж Фарм Фармакол. 2001, 53: 497-503. 10.1211/0022357011775794.
КАС пабмед Статья Google Scholar
Smith QR, Momma S, Aoyagi M, Rapoport SI: Кинетика транспорта нейтральных аминокислот через гематоэнцефалический барьер. Дж. Нейрохим. 1987, 49: 1651-1658. 10.1111/j.1471-4159.1987.tb01039.x.
КАС пабмед Статья Google Scholar
Stoll J, Wadhwani KC, Smith QR: Идентификация переносчика катионных аминокислот (система y+) крысиного гематоэнцефалического барьера. Дж. Нейрохим. 1993, 60: 1956-1959. 10.1111/j.1471-4159.1993.tb13428.x.
Дж. Нейрохим. 1993, 60: 1956-1959. 10.1111/j.1471-4159.1993.tb13428.x.
КАС пабмед Статья Google Scholar
Betz AL, Goldstein GW: Полярность гематоэнцефалического барьера: транспорт нейтральных аминокислот в изолированные капилляры головного мозга. Наука. 1978, 202: 225-226. 10.1126/наук.211586.
КАС пабмед Статья Google Scholar
Tayarani I, Lefauconnier J-M, Roux F, Bourrer J-M: Доказательства наличия аланиновой, сериновой и цистиновой систем транспорта в изолированных капиллярах головного мозга. J Cereb Blood Flow Metab. 1987, 7: 585-591.
КАС пабмед Статья Google Scholar
Санчес дель Пино М.М., Петерсон Д.Р., Хокинс Р.А.: Характеристика транспорта нейтральных аминокислот в изолированных люминальных и аблюминальных мембранах гематоэнцефалического барьера. Дж. Биол. Хим. 1995, 270: 14913-14918.
Дж. Биол. Хим. 1995, 270: 14913-14918.
ПабМед Статья Google Scholar
Lee WJ, Hawkins RA, Viña JR, Peterson DR: Транспорт глутамина через гематоэнцефалический барьер: возможный механизм удаления азота. Am J Physiol Cell Physiol. 1998, 274: C1101-C1107.
КАС Google Scholar
O’Kane RL, Martinez-Lopez I, DeJoseph MR, Viña JR, Hawkins RA: Na(+)-зависимые переносчики глутамата (EAAT1, EAAT2 и EAAT3) гематоэнцефалического барьера.Механизм удаления глутамата. Дж. Биол. Хим. 1999, 274: 31891-31895.
ПабМед Статья Google Scholar
Стегинк Л.Д., Филер Л.Дж., Бейкер Г.Л.: Концентрация аминокислот в плазме у нормальных взрослых, получавших пищу с добавлением L-глутамата натрия и аспартама. Дж Нутр. 1983, 113: 1851-1860.
КАС пабмед Google Scholar

Benveniste H, Drejer J, Schousboe A, Diemer NH: Повышение внеклеточных концентраций глутамата и аспартата в гиппокампе крыс во время транзиторной церебральной ишемии под контролем внутримозгового микродиализа.Дж. Нейрохим. 1984, 43: 1369-1374. 10.1111/j.1471-4159.1984.tb05396.x.
КАС пабмед Статья Google Scholar
Хокинс Р.А.: Гематоэнцефалический барьер и глутамат. Am J Clin Nutr. 2009, 90: 867С-874С. 10.3945/ajcn.2009.27462BB.
Центральный пабмед КАС пабмед Статья Google Scholar
Злотник А., Гуревич Б., Ткачов С., Маоз И., Шапира Ю., Тейхберг В.И.: Нейропротекция мозга путем удаления глутамата крови.Опыт Нейрол. 2007, 203: 213-220. 10.1016/ж.экспнейрол.2006.08.021.
КАС пабмед Статья Google Scholar
Abbott NJ, Patabendige AA, Dolman DE, Yusof SR, Begley DJ: Структура и функция гематоэнцефалического барьера. Нейробиол Дис. 2010, 37: 13-25. 10.1016/j.nbd.2009.07.030.
Нейробиол Дис. 2010, 37: 13-25. 10.1016/j.nbd.2009.07.030.
КАС пабмед Статья Google Scholar
Segal MB, Preston JE, Collis CS, Zlokovic BV: Кинетика и независимость Na от поглощения аминокислот кровью перфузируемого сосудистого сплетения овец.Am J Physiol. 1990, 258: F1288-F1294.
КАС пабмед Google Scholar
Duelli R, Enerson BE, Gerhart DZ, Drewes LR: Экспрессия переносчика больших аминокислот LAT1 в эндотелии мозга крыс. J Cereb Blood Flow Metab. 2000, 20: 1557-1562. 10.1097/00004647-200011000-00005.
КАС пабмед Статья Google Scholar
Preston JE, Segal MB: Поглощение анионных и катионных аминокислот изолированным перфузируемым сосудистым сплетением овец.Мозг Res. 1992, 581: 351-355. 10.1016/0006-8993(92)-Н.
КАС пабмед Статья Google Scholar

Keep RF, Xiang J. Транспорт аминокислот N-системы через гемато-ликворный барьер. Дж. Нейрохим. 1995, 65: 2571-2576. 10.1046/j.1471-4159.1995.65062571.x.
КАС пабмед Статья Google Scholar
Agulhon C, Rostaing P, Ravassard P, Sagné C, Triller A, Giros B: Переносчик лизосомных аминокислот LYAAT-1 в центральной нервной системе крыс: гибридизация in situ и иммуногистохимическое исследование.J Комп Нейрол. 2003, 462: 71-89. 10.1002/cne.10712.
КАС пабмед Статья Google Scholar
Al-Sarraf H, Preston JE, Segal MB: Накопление кислых аминокислот хориоидальным сплетением крыс во время развития. Brain Res Dev Мозг Res. 1997, 102: 47-52. 10.1016/С0165-3806(97)00075-8.
КАС пабмед Статья Google Scholar
Beschorner R, Pantazis G, Jeibmann A, Boy J, Meyermann R, Mittelbronn M, Schittenhelm J: Экспрессия EAAT-1 отличает опухоли сосудистого сплетения от нормального и реактивного эпителия сосудистого сплетения. Акта Нейропатол. 2009, 117: 667-675. 10.1007/с00401-009-0519-у.
Акта Нейропатол. 2009, 117: 667-675. 10.1007/с00401-009-0519-у.
КАС пабмед Статья Google Scholar
Пеллерин Э., Маджистретти П.Дж.: Пища для размышлений: вызов догмам. J Cereb Blood Flow Metab. 2003, 23: 1282-1286. 10.1097/01.ВЦБ.0000096064.12129.3Д.
ПабМед Статья Google Scholar
Adle-Biassette H, Olivier P, Verney C, Fontaine RH, Evrard P, Hénin D, Massias L, Gressens P, Baud O: Кортикальные последствия блокады транспорта монокарбоксилатов in vivo во время развития мозга у мышей.Педиатр Рез. 2007, 61: 54-60. 10.1203/01.пдр.0000250040.61888.61.
КАС пабмед Статья Google Scholar
Poole RC, Halestrap AP: Транспорт лактата и других монокарбоксилатов через плазматические мембраны млекопитающих. Am J Physiol. 1993, 264: C761-C782.
КАС пабмед Google Scholar
Halestrap AP, Meredith D: Семейство генов SLC16: от переносчиков монокарбоксилатов (MCT) до переносчиков ароматических аминокислот и далее.Арка Пфлюгера. 2004, 447: 619-628. 10.1007/s00424-003-1067-2.
КАС пабмед Статья Google Scholar
Бергерсен Л.Х. Является ли лактат пищей для нейронов? Сравнение подтипов переносчиков монокарбоксилатов в головном мозге и мышцах. Неврология. 2007, 145: 11-19. 10.1016/j.neuroscience.2006.11.062.
КАС пабмед Статья Google Scholar
Bergersen L, Wærhaug O, Helm J, Thomas M, Laake P, Davies AJ, Wilson MC, Halestrap AP, Ottersen OP: Новый белок постсинаптической плотности: транспортер монокарбоксилата MCT2 солокализован с дельта-2 глутаматными рецепторами в постсинаптических плотностях параллельных волокна-клетки Пуркинье синапсы. Опыт Мозг Res. 2001, 136: 523-534. 10.1007/s002210000600.
КАС пабмед Статья Google Scholar

Roberts LM, Woodford K, Zhou M, Black DS, Haggerty JE, Tate EH, Grindstaff KK, Mengesha W, Raman C, Zerangue N: Экспрессия переносчиков гормонов щитовидной железы монокарбоксилатного транспортера-8 (SLC16A2) и транспортер органических ионов-14 (SLCO1C1) на гематоэнцефалическом барьере.Эндокринология. 2008, 149: 6251-6261. 10.1210/en.2008-0378.
КАС пабмед Статья Google Scholar
Schwartz CE, Stevenson RE: Переносчик гормонов щитовидной железы MCT8 и синдром Аллана-Херндона-Дадли. Best Pract Res Clin Endocrinol Metab. 2007, 21: 307-321. 10.1016/j.beem.2007.03.009.
Центральный пабмед КАС пабмед Статья Google Scholar
Puchowicz MA, Xu K, Sun X, Ivy A, Emancipator D, LaManna JC: кетоз, вызванный диетой, увеличивает плотность капилляров без изменения кровотока в мозге крыс. Am J Physiol Endocrinol Metab. 2006, 292: E1607-E1615. 10.1152/ажпендо.00512.2006.
10.1152/ажпендо.00512.2006.
Артикул КАС Google Scholar
Лейно Р.Л., Герхарт Д.З., Древес Л.Р.: Обилие переносчиков монокарбоксилатов (MCT1) в головном мозге сосущих и взрослых крыс: количественное электронно-микроскопическое исследование иммунозолота.Brain Res Dev Мозг Res. 1999, 113: 47-54. 10.1016/С0165-3806(98)00188-6.
КАС пабмед Статья Google Scholar
Koehler-Stec EM, Simpson IA, Vannucci SJ, Landschulz KT, Landschulz WH: Экспрессия переносчика монокарбоксилатов в мозге мыши. Am J Physiol. 1998, 275: E516-E524.
КАС пабмед Google Scholar
Permanne B, Adessi C, Saborio GP, Fraga S, Frossard MJ, Van Dorpe J, Dewachter I, Banks WA, Van Leuven F, Soto C: Снижение амилоидной нагрузки и повреждения головного мозга в модели трансгенных мышей болезни Альцгеймера путем лечения пептидом бета-слоя. FASEB J. 2002, 16: 860-862.
FASEB J. 2002, 16: 860-862.
КАС пабмед Google Scholar
Tuszynski MH, Blesch A: Фактор роста нервов: от животных моделей дегенерации холинергических нейронов до генной терапии болезни Альцгеймера. Прог Мозг Res. 2004, 146: 441-449.
КАС пабмед Google Scholar
Доубарн Д., Аллен С.Дж.: Нейротрофины и нейродегенерация.Приложение Нейропатол Нейробиол. 2003, 29: 211-230. 10.1046/j.1365-2990.2003.00487.х.
КАС пабмед Статья Google Scholar
Espejo M, Cutillas B, Arenas TE, Ambrosio S: Увеличение выживаемости дофаминергических нейронов в стриарных трансплантатах вентральных мезэнцефальных клеток плода, подвергшихся воздействию нейротрофина-3 или нейротрофического фактора, полученного из линии глиальных клеток. Трансплантация клеток. 2000, 9: 45-53.
КАС пабмед Google Scholar

Guan J, Krishnamurthi R, Waldvogel HJ, Faull RL, Clark R, Gluckman P: N-концевой трипептид IGF-1 (GPE) предотвращает потерю TH-позитивных нейронов после индуцированного 6-OHDA поражения ниграла у крыс. Мозг Res. 2000, 859: 286-292. 10.1016/S0006-8993(00)01988-0.
КАС пабмед Статья Google Scholar
Brasnjevic I, Steinbusch HW, Schmitz C, Martinez-Martinez P: Доставка пептидных и белковых препаратов через гематоэнцефалический барьер.Прог Нейробиол. 2009, 87: 212-251. 10.1016/ж.пневробио.2008.12.002.
КАС пабмед Статья Google Scholar
Zlokovic BV, Begley D, Chain DG: Проницаемость гематоэнцефалического барьера для дипептидов и составляющих их аминокислот. Мозг Res. 1983, 271: 65-71. 10.1016/0006-8993(83)
КАС пабмед Статья Google Scholar
Rubio-Aliaga I, Daniel H: Переносчики пептидов и их роль в физиологических процессах и распределении лекарств. Ксенобиотика. 2008, 38: 1022-1042. 10.1080/00498250701875254.
Ксенобиотика. 2008, 38: 1022-1042. 10.1080/00498250701875254.
КАС пабмед Статья Google Scholar
Zlokovic BV, Hyman S, McComb JG, Lipovac MN, Tang G, Davson H: Кинетика поглощения аргинин-вазопрессина через гематоэнцефалический барьер. Биохим Биофиз Акта. 1990, 1025: 191-198. 10.1016/0005-2736(90)-8.
КАС пабмед Статья Google Scholar
Zlokovic BV, Lipovac MN, Begley DJ, Davson H, Rakic L: Транспорт лейцин-энкефалина через гематоэнцефалический барьер в перфузированном мозге морской свинки. Дж. Нейрохим. 1987, 49: 310-315. 10.1111/j.1471-4159.1987.tb03431.x.
КАС пабмед Статья Google Scholar
Zlokovic BV, Mackic JB, Djuricic B, Davson H: Кинетический анализ поглощения лейцин-энкефалина клетками на просветной стороне гематоэнцефалического барьера в перфузированном in situ мозге морской свинки. Дж. Нейрохим. 1989, 53: 1333-1340. 10.1111/j.1471-4159.1989.tb08522.x.
Дж. Нейрохим. 1989, 53: 1333-1340. 10.1111/j.1471-4159.1989.tb08522.x.
КАС пабмед Статья Google Scholar
Злокович Б.В. Проницаемость сосудов головного мозга для пептидов: манипуляции с транспортными системами на гематоэнцефалическом барьере. Фарм Рез. 1995, 12: 1395-1406. 10.1023/А:1016254514167.
КАС пабмед Статья Google Scholar
Banks WA, Kastin AJ: Транспортная система, опосредованная переносчиком из мозга в кровь, для небольших N-тирозинированных пептидов. Фармакол Биохим Поведение. 1984, 21: 943-946. 10.1016/С0091-3057(84)80077-5.
КАС пабмед Статья Google Scholar
Banks WA: ЦНС как мишень для пептидов и препаратов на их основе. Экспертное заключение Препарат Делив. 2006, 3: 707-712. 10.1517/17425247.3.6.707.
КАС пабмед Статья Google Scholar
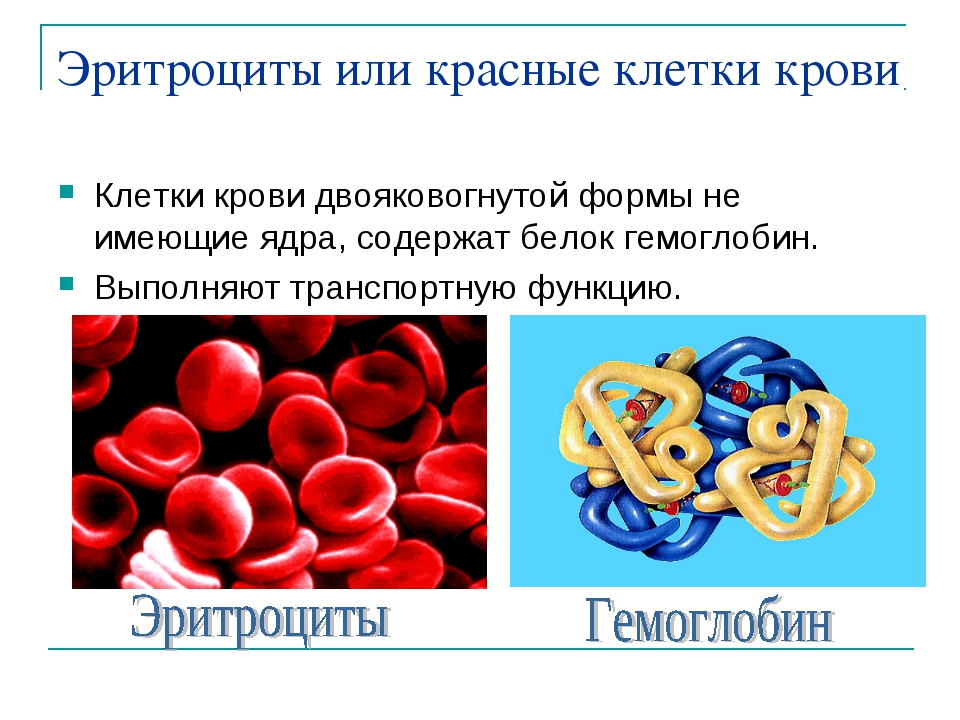
Dogrukol-Ak D, Kumar VB, Ryerse JS, Farr SA, Verma S, Nonaka N, Nakamachi T, Ohtaki H, Niehoff ML, Edwards JC, Shioda S, Morley JE, Banks WA: Выделение пептидной транспортной системы-6 из эндотелиальные клетки головного мозга: терапевтические эффекты антисмыслового ингибирования в моделях болезни Альцгеймера и инсульта. J Cereb Blood Flow Metab. 2009, 29: 411-422. 10.1038/jcbfm.2008.131.
КАС пабмед Статья Google Scholar
Brownlees J, Williams CH: Пептидазы, пептиды и гематоэнцефалический барьер млекопитающих.Дж. Нейрохим. 1993, 60: 793-803. 10.1111/j.1471-4159.1993.tb03223.x.
КАС пабмед Статья Google Scholar
Бэнкс В. А., Кастин А. Дж.: Взаимоотношения ЦСЖ-плазма для DSIP и некоторых других нейропептидов. Фармакол Биохим Поведение. 1983, 19: 1037-1040. 10.1016/0091-3057(83)
-4.КАС пабмед Статья Google Scholar
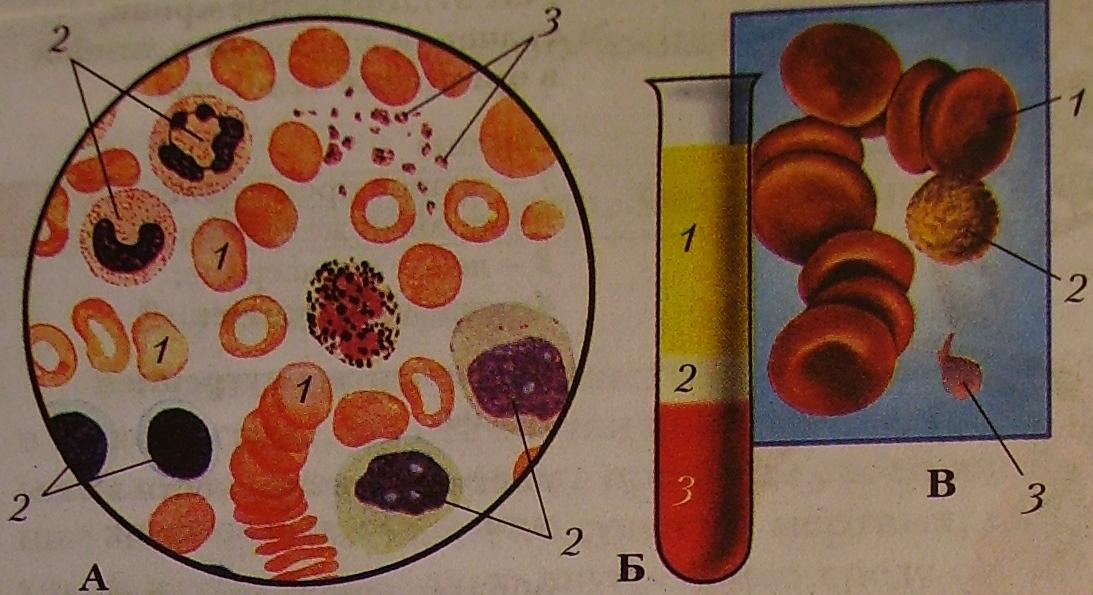
Woods SC, Seeley RJ, Baskin DG, Schwartz MW: Инсулин и гематоэнцефалический барьер.Курр Фарм Дез. 2003, 9: 795-800. 10.2174/1381612033455323.
КАС пабмед Статья Google Scholar
Broadwell RD, Baker-Cairns BJ, Friden PM, Oliver C, Villegas JC: Трансцитоз белка через эпителий и эндотелий головного мозга млекопитающих. III. Рецептор-опосредованный трансцитоз через гематоэнцефалический барьер переносимого кровью трансферрина и антител против рецептора трансферрина. Опыт Нейрол. 1996, 142: 47-65.10.1006/доб.1996.0178.
КАС пабмед Статья Google Scholar
Banks WA, Farr SA, La Scola ME, Morley JE: Внутривенное введение человеческого интерлейкина-1альфа ухудшает обработку памяти у мышей: зависимость от транспорта через гематоэнцефалический барьер в задний отдел перегородки. J Pharmacol Exp Ther. 2001, 299: 536-541.
КАС пабмед Google Scholar

Злокович Б.В., Йованович С., Мяо В., Самара С., Верма С., Фаррелл К.Л.: Дифференциальная регуляция транспорта лептина сосудистым сплетением и гематоэнцефалическим барьером и высокоаффинными транспортными системами для входа в гипоталамус и через гемато-спинномозговой барьер . Эндокринология. 2000, 141: 1434-1441. 10.1210/ан.141.4.1434.
КАС пабмед Google Scholar
Banks WA, Farrell CL: Нарушение транспорта лептина через гематоэнцефалический барьер при ожирении является приобретенным и обратимым.Am J Physiol Endocrinol Metab. 2003, 285: Е10-Е15.
КАС пабмед Статья Google Scholar
Deane R, Sagare A, Hamm K, Parisi M, LaRue B, Guo H, Wu Z, Holtzman DM, Zlokovic BV: IgG-зависимый клиренс бета-амилоидного пептида болезни Альцгеймера кровью и мозгом барьерный неонатальный Fc-рецептор. Дж. Нейроски. 2005, 25: 11495-11503. 10.1523/JNEUROSCI. 3697-05.2005.
3697-05.2005.
КАС пабмед Статья Google Scholar
Нисидзима Т., Пириз Дж., Дюфлот С., Фернандес А.М., Гайтан Г., Гомес-Пинедо У., Вердуго Дж.М., Лерой Ф., Соя Х., Нуньес А., Торрес-Алеман И.: Нейрональная активность управляет локальным гематоэнцефалическим барьерным транспортом сыворотки инсулиноподобного фактора роста-I в ЦНС. Нейрон. 2010, 67: 834-846. 10.1016/j.neuron.2010.08.007.
КАС пабмед Статья Google Scholar
Biere AL, Ostaszewski B, Stimson ER, Hyman BT, Maggio JE, Selkoe DJ: Амилоидный бета-пептид транспортируется липопротеинами и альбумином в плазме человека.Дж. Биол. Хим. 1996, 271: 32916-32922. 10.1074/jbc.271.51.32916.
КАС пабмед Статья Google Scholar
Calero M, Rostagno A, Matsubara E, Zlokovic B, Frangione B, Ghiso J: Аполипопротеин J (кластерин) и болезнь Альцгеймера. Микроск Рес Тех. 2000, 50: 305-315. 10.1002/1097-0029(20000815)50:4<305::AID-JEMT10>3.0.CO;2-л.
Микроск Рес Тех. 2000, 50: 305-315. 10.1002/1097-0029(20000815)50:4<305::AID-JEMT10>3.0.CO;2-л.
КАС пабмед Статья Google Scholar
Сагаре А., Дин Р., Белл Р.Д., Джонсон Б., Хамм К., Пенду Р., Марки А., Лентинг П.Дж., Ву З., Зарконе Т., Коза А., Майо К., Перлмуттер Д., Кома М., Чжун З., Злокович Б.В.: Допуск бета-амилоида циркулирующими рецепторами липопротеинов. Нат Мед. 2007, 13: 1029-1031. 10,1038/нм1635.
Центральный пабмед КАС пабмед Статья Google Scholar
Deane R, Bell RD, Sagare A, Zlokovic BV: Клиренс амилоид-бета-пептида через гематоэнцефалический барьер: значение для лечения болезни Альцгеймера.Лекарственные мишени для нейролептиков ЦНС. 2009, 8: 16-30. 10.2174/187152709787601867.
Центральный пабмед КАС пабмед Статья Google Scholar

Ян С.Ф., Равичандран Р., Шмидт А.М.: Ось RAGE: основной механизм, сигнализирующий об опасности для уязвимой сосудистой системы. Цирк Рез. 2010, 106: 842-853. 10.1161/ЦИРКРЕСАХА.109.212217.
Центральный пабмед КАС пабмед Статья Google Scholar
Злокович Б.В., Дин Р., Сагаре А.П., Белл Р.Д., Винклер Э.А. Белок-1, родственный рецептору липопротеинов низкой плотности: гомеостатический механизм серийного клиренса, контролирующий элиминацию амилоидного β-пептида болезни Альцгеймера из головного мозга. Дж. Нейрохим. 2010, 115: 1077-1089. 10.1111/j.1471-4159.2010.07002.х.
Центральный пабмед КАС пабмед Статья Google Scholar
Дин Р., Ву З., Сагаре А., Дэвис Дж., Ду Янь С., Хамм К., Сюй Ф., Паризи М., ЛаРю Б., Ху Х.В., Спийкерс П., Го Х., Сонг Х., Лентинг П.Дж., Ван Ностранд В.Е., Злокович Б.В.: взаимодействие LRP/амилоидного b-пептида опосредует дифференциальный отток изоформ антител в головной мозг. Нейрон. 2004, 43: 333-344. 10.1016/j.neuron.2004.07.017.
Нейрон. 2004, 43: 333-344. 10.1016/j.neuron.2004.07.017.
КАС пабмед Статья Google Scholar
Белл Р.Д., Сагаре А.П., Фридман А.Е., Беди Г.С., Хольцман Д.М., Дин Р., Злокович Б.В.: Транспортные пути для клиренса человеческого бета-амилоида Альцгеймера и аполипопротеинов E и J в центральной нервной системе мыши. J Cereb Blood Flow Metab. 2007, 27: 909-918.
Центральный пабмед КАС пабмед Google Scholar
Ito S, Ueno T, Ohtsuki S, Terasaki T: Отсутствие транспортной активности оттока из мозга в кровь белка-1, связанного с рецептором липопротеинов низкой плотности (LRP-1), для амилоида-b-пептида (1-40) в мышь: вовлечение LRP-1-независимого пути. Дж. Нейрохим. 2010, 113: 1356-1363.
КАС пабмед Google Scholar
Banks WA: Доставка пептидов в мозг: акцент на терапевтических разработках. Биополимеры. 2008, 90: 589-594.10.1002/бип.20980.
Биополимеры. 2008, 90: 589-594.10.1002/бип.20980.
КАС пабмед Статья Google Scholar
Broadwell RD: Трансцитоз макромолекул через гематоэнцефалический барьер: клеточная биологическая перспектива и критическая оценка. Акта Нейропатол. 1989, 79: 117-128. 10.1007/BF00294368.
КАС пабмед Статья Google Scholar
Бодор Н., Бухвальд П.: Доставка лекарств в мозг: опыт на сегодняшний день.Am J Drug Deliv. 2003, 1: 13-26. 10.2165/00137696-200301010-00002.
Артикул Google Scholar
Левин В.А. Зависимость коэффициента распределения октанол/вода и молекулярной массы от проницаемости капилляров головного мозга крысы. J Med Chem. 1980, 23: 682-684. 10.1021/jm00180a022.
КАС пабмед Статья Google Scholar
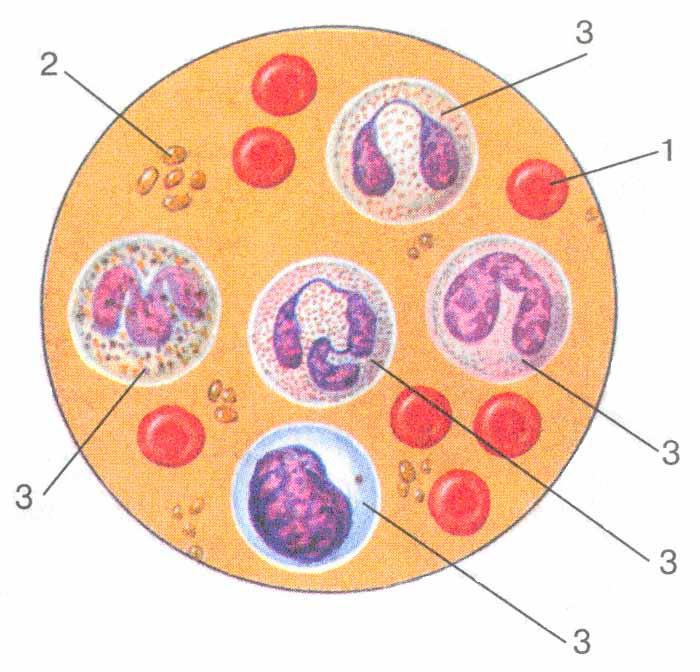
Banks WA, Kastin AJ: Редакционный обзор: Пептидные транспортные системы для опиатов через гематоэнцефалический барьер.Am J Physiol. 1990, 259: Е1-Е10.
КАС пабмед Google Scholar
Кинг М., Су В., Чанг А., Цукерман А., Пастернак Г.В.: Транспорт опиоидов из головного мозга на периферию с помощью Р-гликопротеина: периферические действия центральных препаратов. Нат Нейроски. 2001, 4: 221-222. 10.1038/85115.
Артикул Google Scholar
Boado RJ, Zhang Y, Wang Y, Pardridge WM: Разработка и экспрессия моноклонального антитела к химерному рецептору трансферрина для доставки через гематоэнцефалический барьер у мышей.Биотехнология Биоинж. 2009, 102: 1251-1258. 10.1002/бит.22135.
Центральный пабмед КАС пабмед Статья Google Scholar
van Rooy I, Cakir-Tascioglu S, Couraud PO, Romero IA, Weksler B, Storm G, Hennink WE, Schiffelers RM, Mastrobattista E: Идентификация пептидных лигандов для нацеливания на гематоэнцефалический барьер. Фарм Рез. 2010, 27: 673-682. 10.1007/с11095-010-0053-6.
Фарм Рез. 2010, 27: 673-682. 10.1007/с11095-010-0053-6.
Центральный пабмед КАС пабмед Статья Google Scholar
Шмидынгер-Ходобска Дж., Фокс Л.М., Линч К.М., Цинк Б.Дж., Ходобски А.: Вазопрессин усиливает выработку провоспалительных медиаторов при черепно-мозговой травме. J Нейротравма. 2010, 27: 1449-1461. 10.1089/neu.2010.1331.
Центральный пабмед пабмед Статья Google Scholar
Ocheltree SM, Shen H, Hu Y, Xiang J, Keep RF, Smith DE: Роль PEPT2 в поглощении глицилсаркозина и 5-аминолевулиновой кислоты сосудистым сплетением: исследования на мышах дикого типа и нулевых мышах.Фарм Рез. 2004, 21: 1680-1685. 10.1023/Б:ФАМ.0000041465.89254.05.
КАС пабмед Статья Google Scholar
Yamashita T, Shimada S, Guo W, Sato K, Kohmura E, Hayakawa T, Takagi T, Tohyama M: Клонирование и функциональная экспрессия переносчика пептидов/гистидина в головном мозге. Дж. Биол. Хим. 1997, 272: 10205-10211. 10.1074/jbc.272.17.11408.
Дж. Биол. Хим. 1997, 272: 10205-10211. 10.1074/jbc.272.17.11408.
КАС пабмед Статья Google Scholar
Ocheltree SM, Shen H, Hu Y, Keep RF, Smith DE: Роль и значение переносчика пептидов 2 (PEPT2) в почках и сосудистом сплетении: исследования in vivo с глицилсаркозином у мышей дикого типа и мышей с нокаутом PEPT2. J Pharmacol Exp Ther. 2005, 315: 240-247. 10.1124/жпет.105.089359.
КАС пабмед Статья Google Scholar
Shu C, Shen H, Teuscher NS, Lorenzi PJ, Keep RF, Smith DE: Роль PEPT2 в переносе пептидов/миметиков через гемато-спинномозговой барьер: исследования эпителиальных клеток сосудистого сплетения крыс в первичной культуре .J Pharmacol Exp Ther. 2002, 301: 820-829. 10.1124/jpet.301.3.820.
КАС пабмед Статья Google Scholar
Berger UV, Hediger MA: Распределение мРНК переносчика пептидов PEPT2 в нервной системе крыс. Анат Эмбриол (Берл). 1999, 199: 439-449. 10.1007/s0042
Анат Эмбриол (Берл). 1999, 199: 439-449. 10.1007/s0042
242.
КАС Статья Google Scholar
Dietrich MO, Spuch C, Antequera D, Rodal I, de Yébenes JG, Molina JA, Bermejo F, Carro E: Мегалин опосредует транспорт лептина через гематоэнцефалический барьер.Нейробиол Старение. 2008, 29: 902-912. 10.1016/j.нейробиология.2007.01.008.
КАС пабмед Статья Google Scholar
Alvira-Botero X, Carro EM: Клиренс амилоидного пептида через сосудистое сплетение при болезни Альцгеймера. Curr Старение Sci. 2010.
Google Scholar
Behl M, Zhang Y, Shi Y, Cheng J, Du Y, Zheng W: Индуцированное свинцом накопление бета-амилоида в сосудистом сплетении: роль белка-1 рецептора липопротеинов низкой плотности и протеинкиназы C .Нейротоксикология. 2010, 31: 524-532. 10.1016/ж.нейро.2010.05.004.
Центральный пабмед КАС пабмед Статья Google Scholar

Johanson CE, Duncan JA, Klinge PM, Brinker T, Stopa EG, Silverberg GD: Множественность функций спинномозговой жидкости: новые проблемы в области здоровья и болезней. Цереброспинальная жидкость Res. 2008, 5: 10-10.1186/1743-8454-5-10.
Центральный пабмед пабмед Статья КАС Google Scholar
Johanson CE, Donahue JE, Spangenberger A, Stopa EG, Duncan JA, Sharma HS: Предсердный натрийуретический пептид: его предполагаемая роль в модуляции системы сосудистое сплетение-CSF для регулирования внутричерепного давления. Acta Neurochir Suppl. 2006, 96: 451-456. полный текст.
КАС пабмед Статья Google Scholar
Мори К., Цуцуми К., Курихара М., Кавагути Т., Нива М.: Изменение предсердных рецепторов натрийуретического пептида в сосудистом сплетении крыс с индуцированной или врожденной гидроцефалией.Чайлдс Нерв Сист. 1990, 6: 190-193. 10.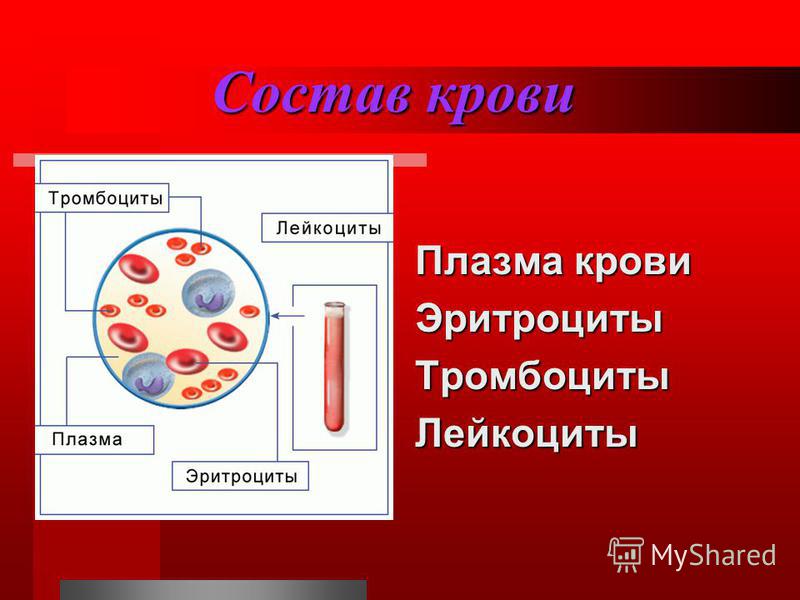 1007/BF01850969.
1007/BF01850969.
КАС пабмед Статья Google Scholar
Chodobski A, Loh YP, Corsetti S, Szmydynger-Chodobska J, Johanson CE, Lim YP, Monfils PR: Наличие аргинин-вазопрессина и его мРНК в эпителии сосудистого сплетения крыс. Мозг Res Мол Мозг Res. 1997, 48: 67-72. 10.1016/S0169-328X(97)00079-X.
КАС пабмед Статья Google Scholar
Szmydynger-Chodobska J, Chung I, Chodobski A: Хроническая гипернатриемия увеличивает экспрессию вазопрессина и потенциалзависимых натриевых каналов в сосудистом сплетении крыс. Нейроэндокринология. 2006, 84: 339-345.
КАС пабмед Google Scholar
Chodobski A, Szmydynger-Chodobska J, Johanson CE: Вазопрессин опосредует ингибирующее действие центрального ангиотензина II на образование спинномозговой жидкости. Евр Дж Фармакол. 1998, 347: 205-209. 10.1016/С0014-2999(98)00229-5.
10.1016/С0014-2999(98)00229-5.
КАС пабмед Статья Google Scholar
Johanson CE, Preston JE, Chodobski A, Stopa EG, Szmydynger-Chodobska J, McMillan PN: Опосредованное рецептором AVP V1 снижение оттока Cl- и увеличение количества темных клеток в эпителии сосудистого сплетения. Am J Physiol. 1999, 276: C82-C90.
КАС пабмед Google Scholar
Johanson C, Stopa E, Baird A, Sharma H: черепно-мозговая травма и механизмы восстановления: пептидная модуляция перивентрикулярных нейрогенных областей сосудистым сплетением-ликворной связью. J Neural Transm. 2010.
Google Scholar
Schreiber G: Эволюция синтеза транстиретина в сосудистом сплетении. Clin Chem Lab Med. 2002, 40: 1200-1210. 10.1515/CCLM.2002.210.
КАС пабмед Статья Google Scholar
Palha JA, Hays MT, de Escobar GM, Episkopou V, Gottesman ME, Saraiva MHM: транстиретин не является необходимым для достижения тироксином мозга и других тканей у транстиретин-нулевых мышей. Am J Physiol. 1997, 272: Е485-Е493.
КАС пабмед Google Scholar
Quintela T, Alves CH, Goncalves I, Baltazar G, Saraiva MJ, Santos CR: 5-альфа-дигидротестостерон повышает уровень транстиретина в хориоидальных сплетениях мышей и крыс посредством пути, независимого от рецептора андрогена.Мозг Res. 2008, 1229: 18-26. 10.1016/j.brainres.2008.06.095.
КАС пабмед Статья Google Scholar
Tang YP, Haslam SZ, Conrad SE, Sisk CL: Эстроген увеличивает экспрессию в мозгу мРНК, кодирующей транстиретин, белок-нейтрализатор бета-амилоида. Дж. Альцгеймера Дис. 2004, 6: 413-420.
КАС пабмед Google Scholar

Quintela T, Goncalves I, Baltazar G, Alves CH, Saraiva MJ, Santos CR: 17бета-эстрадиол индуцирует экспрессию транстиретина в сосудистом сплетении мышей через путь, зависимый от рецептора эстрогена.Селл Мол Нейробиол. 2009, 29: 475-483. 10.1007/s10571-008-9339-1.
КАС пабмед Статья Google Scholar
Quintela T, Goncalves I, Martinho A, Alves CH, Saraiva MJ, Rocha P, Santos CR: Прогестерон усиливает экспрессию транстиретина в сосудистых сплетениях крыс in vitro и in vivo через рецептор прогестерона. Джей Мол Нейроски. 2010.
Google Scholar
Schinkel AH, Jonker JW: переносчики оттока лекарств из млекопитающих семейства кассет связывания АТФ (ABC): обзор:. Adv Drug Deliv Rev. 2003, 55: 3-29. 10.1016/S0169-409X(02)00169-2.
КАС пабмед Статья Google Scholar
Сауна З. Е., Смит М.М., Мюллер М., Керр К.М., Амбудкар С.М.: Механизм действия связанного с множественной лекарственной устойчивостью Р-гликопротеина. J Биоэнергетические биомембраны. 2001, 33: 481-491. 10.1023/А:1012875105006.
Е., Смит М.М., Мюллер М., Керр К.М., Амбудкар С.М.: Механизм действия связанного с множественной лекарственной устойчивостью Р-гликопротеина. J Биоэнергетические биомембраны. 2001, 33: 481-491. 10.1023/А:1012875105006.
КАС Статья Google Scholar
Russel FG, Masereeuw R, van Aubel RA: Молекулярные аспекты почечного транспорта анионных препаратов. Энн Рев Физиол. 2002, 64: 563-594. 10.1146/аннурев.физиол.64.081501.155913.
КАС Статья Google Scholar
Dresser MJ, Leabman MK, Giacomini KM: Транспортеры, участвующие в выведении лекарств в почках: переносчики органических анионов и переносчики органических катионов.Дж. Фарм. 2001, 90: 397-421. 10.1002/1520-6017(200104)90:4<397::AID-JPS1000>3.0.CO;2-D.
КАС пабмед Статья Google Scholar
Roberts LM, Black DS, Raman C, Woodford K, Zhou M, Haggerty JE, Yan AT, Cwirla SE, Grindstaff KK: Субклеточная локализация транспортеров вдоль гематоэнцефалического барьера крысы и гематоэнцефалического барьера жидкий барьер путем биотинилирования in vivo. Неврология. 2008, 155: 423-438.10.1016/j.neuroscience.2008.06.015.
Неврология. 2008, 155: 423-438.10.1016/j.neuroscience.2008.06.015.
КАС пабмед Статья Google Scholar
Бабаханян К., Бендаян М., Бендаян Р.: Локализация Р-гликопротеина в ядерной оболочке клеток головного мозга крысы. Biochem Biophys Res Commun. 2007, 361: 301-306. 10.1016/j.bbrc.2007.06.176.
КАС пабмед Статья Google Scholar
Schinkel A, Mayer U, Wagenaar E, Mol C, van Deemter L, Smit J, van der Valk MA, Voordouw AC, Spits H, van Tellingen O, Zijlmans JM, Fibbe WE, Borst P: нормальный жизнеспособность и измененная фармакокинетика у мышей, лишенных Р-гликопротеинов типа mdr1 (переносящих лекарство).Proc Natl Acad Sci USA. 1997, 94: 4028-4033. 10.1073/пнас.94.8.4028.
Центральный пабмед КАС пабмед Статья Google Scholar
Linnet K, Ejsing T: Обзор влияния P-гликопротеина на проникновение лекарств в мозг. Сосредоточьтесь на психотропных препаратах. Евро Нейропсихофармакол. 2008, 18: 157-169. 10.1016/j.euroneuro.2007.06.003.
Сосредоточьтесь на психотропных препаратах. Евро Нейропсихофармакол. 2008, 18: 157-169. 10.1016/j.euroneuro.2007.06.003.
КАС пабмед Статья Google Scholar
Miller DS: Регуляция P-гликопротеина и других переносчиков лекарств ABC через гематоэнцефалический барьер. Trends Pharmacol Sci. 2010, 31: 246-54. 10.1016/j.tips.2010.03.003.
Центральный пабмед КАС пабмед Статья Google Scholar
Eyal S, Hsiao P, Unadkat JD: Лекарственные взаимодействия через гематоэнцефалический барьер: факт или фантазия?. Фармакол Тер. 2009, 123: 80-104. 10.1016/j.pharmthera.2009.03.017.
Центральный пабмед КАС пабмед Статья Google Scholar
Calatozzolo C, Gelati M, Ciusani E, Sciacca F, Pollo B, Cajola L, Marras C, Silvani A, Vitellaro-Zuccarello L, Croci D, Boiardi A, Salmaggi A: Экспрессия белков лекарственной устойчивости Pgp, MRP1, MRP3, MRP5 и GST-pi в глиоме человека. Дж. Нейроонкол. 2005, 74: 113-121. 10.1007/s11060-004-6152-7.
Дж. Нейроонкол. 2005, 74: 113-121. 10.1007/s11060-004-6152-7.
КАС пабмед Статья Google Scholar
Чжан В., Мойсилович-Петрович Дж., Андраде М., Чжан Х., Болл М., Станимирович Д.: Экспрессия и функциональная характеристика ABCG2 в эндотелиальных клетках и сосудах головного мозга.FASEB J. 2003, 17: 2085-2087.
ПабМед Google Scholar
Dauchy S, Dutheil F, Weaver R, Chassoux F, Daumas-Duport C, Couraud P, Sherrmann JM, De Waziers I, Declèves X: Транспортеры ABC, цитохромы P450 и их основные факторы транскрипции: экспрессия в гематоэнцефалический барьер человека. Дж. Нейрохим. 2008, 107: 1518-1528. 10.1111/j.1471-4159.2008.05720.х.
КАС пабмед Статья Google Scholar
Westholm D, Rumbley J, Salo D, Rich T, Anderson G: Полипептиды, транспортирующие органические анионы, на гематоэнцефалическом и гематоэнцефалическом барьерах. Curr Top Dev Biol. 2008, 80: 135-170. полный текст.
Curr Top Dev Biol. 2008, 80: 135-170. полный текст.
КАС пабмед Статья Google Scholar
Bronger H, König J, Kopplow K, Steiner H, Ahmadi R, Herold-Mende C, Keppler D, Nies AT: ABCC насосы для оттока лекарств и транспортеры захвата органических анионов в глиомах человека и гематоопухолевый барьер .Рак рез. 2005, 65: 11419-11428. 10.1158/0008-5472.CAN-05-1271.
КАС пабмед Статья Google Scholar
Mori S, Ohtsuki S, Takanaga H, Kikkawa T, Kang Y, Terasaki T: Переносчик органических анионов 3 участвует в оттоке из мозга в кровь аналогов тиопуриновых азотистых оснований. Дж. Нейрохим. 2004, 90: 931-941. 10.1111/j.1471-4159.2004.02552.х.
КАС пабмед Статья Google Scholar
Alebouyeh M, Takeda M, Onozato M, Tojo A, Noshiro R, Hasannejad H, Inatomi J, Narikawa S, Huang XL, Khamdang S, Anzai N, Endou H: экспрессия переносчиков органических анионов человека в сосудистом сплетении и их взаимодействие с метаболитами нейромедиаторов. J Pharmacol Sci. 2003, 93: 430-436. 10.1254/jphs.93.430.
J Pharmacol Sci. 2003, 93: 430-436. 10.1254/jphs.93.430.
КАС пабмед Статья Google Scholar
Koepsell H, Lips K, Volk C: Полиспецифические переносчики органических катионов: структура, функция, физиологические роли и биофармацевтические последствия.Фарм Рез. 2007, 24: 1227-1251. 10.1007/с11095-007-9254-з.
КАС пабмед Статья Google Scholar
Miecz D, Januszewicz E, Czeredys M, Hinton B, Berezowski V, Cecchelli R, Nałecz KA: Локализация переносчика органических катионов/карнитина (OCTN2) в клетках, образующих гематоэнцефалический барьер. Дж. Нейрохим. 2008, 104: 113-123.
КАС пабмед Google Scholar
Strazielle N, Khuth ST, Ghersi-Egea JF: Системы детоксикации, пассивный и специфический транспорт лекарств через гематоэнцефалический барьер в нормальных и патологических ситуациях. Adv Drug Deliv Rev. 2004, 56: 1717-1740. 10.1016/j.addr.2004.07.006.
Adv Drug Deliv Rev. 2004, 56: 1717-1740. 10.1016/j.addr.2004.07.006.
КАС пабмед Статья Google Scholar
Daood M, Tsai C, Ahdab-Barmada M, Watchko J: ABC Transporter (P-gp/ABCB1, MRP1/ABCC1, BCRP/ABCG2) Экспрессия в развивающейся ЦНС человека.нейропедиатрия. 2008, 39: 211-218. 10.1055/с-0028-1103272.
Центральный пабмед КАС пабмед Статья Google Scholar
Rao VV, Dahlheimer JL, Bardgett ME, Snyder AZ, Finch RA, Sartorelli AC, Piwnica-Worms D: Эпителиальная экспрессия гликопротеина P MDR1 P и белка, ассоциированного с множественной лекарственной устойчивостью, в эпителии сосудистого жидкий лекарственно-проницаемый барьер. Proc Natl Acad Sci USA. 1999, 96: 3900-3905.10.1073/пнас.96.7.3900.
Центральный пабмед КАС пабмед Статья Google Scholar
Gazzin S, Strazielle N, Schmitt C, Fevre-Montange M, Ostrow JD, Tiribelli C, Ghersi-Egea JF: Дифференциальная экспрессия белков ABCb1 и ABCc1, связанных с множественной лекарственной устойчивостью, между кровью и мозгом. J Комп Нейрол. 2008, 510: 497-507. 10.1002/cne.21808.
J Комп Нейрол. 2008, 510: 497-507. 10.1002/cne.21808.
КАС пабмед Статья Google Scholar
Nies A, Jedlitschky G, König J, Herold-Mende C, Steiner H, Schmitt H, Keppler D: Экспрессия и иммунолокализация белков множественной лекарственной устойчивости MRP1-MRP6 (ABCC1-ABCC6) в головном мозге человека. Неврология. 2004, 129: 349-360. 10.1016/j.neuroscience.2004.07.051.
КАС пабмед Статья Google Scholar
Lee YJ, Kusuhara H, Sugiyama Y: Играют ли белки-1 и -2, ассоциированные с множественной лекарственной устойчивостью, какую-либо роль в элиминации бета-глюкуронида эстрадиола-17 и 2,4-динитрофенил-S-глутатиона через барьер кровь-цереброспинальная жидкость?.Дж. Фарм. 2004, 93: 99-107. 10.1002/jps.10521.
КАС пабмед Статья Google Scholar
Тачикава М., Ватанабэ М. , Хори С., Фукая М., Оцуки С., Асасима Т., Терасаки Т.: Различная пространственно-временная экспрессия транспортеров ABCA и ABCG в развивающемся и взрослом мозге мыши. Дж. Нейрохим. 2005, 95: 294-304. 10.1111/j.1471-4159.2005.03369.х.
, Хори С., Фукая М., Оцуки С., Асасима Т., Терасаки Т.: Различная пространственно-временная экспрессия транспортеров ABCA и ABCG в развивающемся и взрослом мозге мыши. Дж. Нейрохим. 2005, 95: 294-304. 10.1111/j.1471-4159.2005.03369.х.
КАС пабмед Статья Google Scholar
Gao B, Stieger B, Noe B, Fritschy JM, Meier PJ: Локализация полипептида 2, транспортирующего органические анионы (Oatp2), в эндотелии капилляров и эпителии сосудистого сплетения головного мозга крыс. J Гистохим Цитохим. 1999, 47: 1255-1264.
КАС пабмед Статья Google Scholar
Kusuhara H, He Z, Nagata Y, Nozaki Y, Ito T, Masuda H, Meier PJ, Abe T, Sugiyama Y: Экспрессия и функциональное участие полипептида, транспортирующего органические анионы, подтипа 3 (Slc21a7) в сосудистой оболочке крысы сплетение.Фарм Рез. 2003, 20: 720-727. 10.1023/А:1023473216759.
КАС пабмед Статья Google Scholar
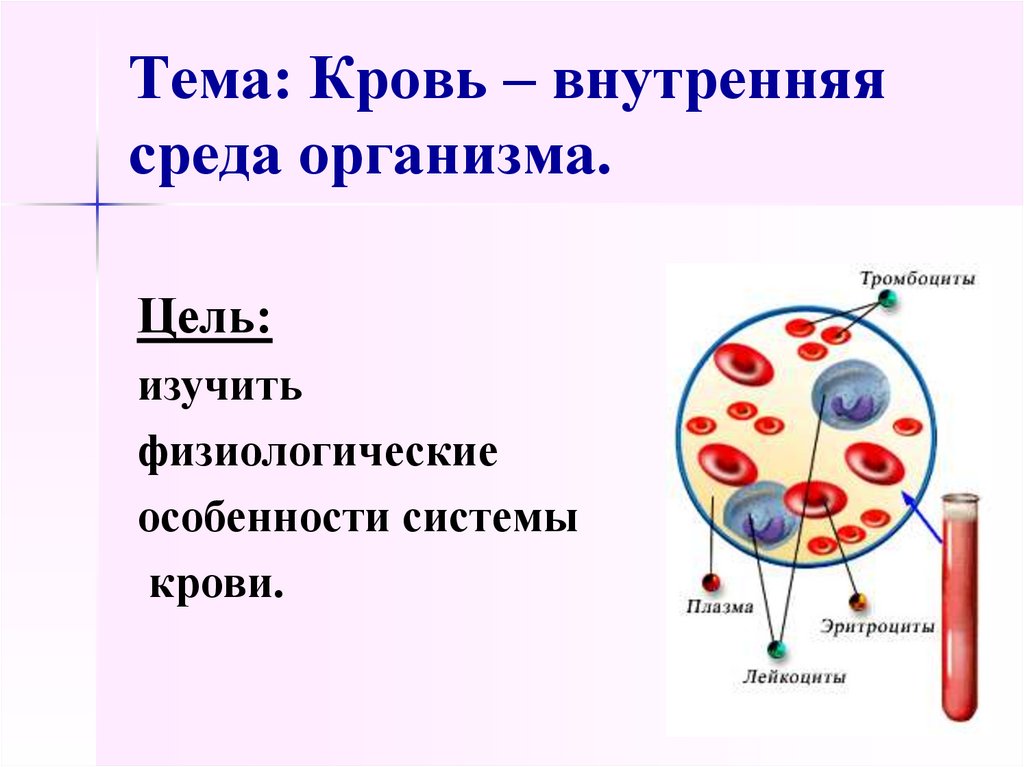
Zhang H, Song YN, Liu WG, Guo XL, Yu LG: Регуляция и роль полипептидов, транспортирующих органические анионы (OATP), в доставке лекарств в сосудистое сплетение. Дж. Клин Нейроски. 2010, 17: 679-684. 10.1016/j.jocn.2009.11.001.
КАС пабмед Статья Google Scholar
Чоудхури С., Черрингтон Н.Дж., Ли Н., Клаассен К.Д.: Конститутивная экспрессия различных мРНК переносчиков ксенобиотиков и эндобиотиков в сосудистых сплетениях крыс. Препарат Метаб Распоряжение. 2003, 31: 1337-1345. 10.1124/дмд.31.11.1337.
КАС пабмед Статья Google Scholar
Nagata Y, Kusuhara H, Endou H, Sugiyama Y: Экспрессия и функциональная характеристика крысиного переносчика органических анионов 3 (rOat3) в сосудистом сплетении.Мол Фармакол. 2002, 61: 982-988. 10.1124/моль.61.5.982.
КАС пабмед Статья Google Scholar

Bauer B, Hartz AM, Fricker G, Miller DS: Pregnane × рецепторная активация экспрессии P-гликопротеина и транспортной функции через гематоэнцефалический барьер. Мол Фармакол. 2004, 66: 413-419.
КАС пабмед Google Scholar
Bauer B, Yang X, Hartz AM, Olson ER, Zhao R, Kalvass JC, Pollack GM, Miller DS: In vivo активация человеческого рецептора прегнана × укрепляет гематоэнцефалический барьер для метадона через P-гликопротеин повышающая регуляция.Мол Фармакол. 2006, 70: 1212-1219. 10.1124/моль.106.023796.
КАС пабмед Статья Google Scholar
Roberts DJ, Goralski KB: Критический обзор влияния воспаления и инфекции на экспрессию и активность P-гликопротеина в головном мозге. Мнение эксперта Препарат Метаб Токсикол. 2008, 4: 1245-1264. 10.1517/17425255.4.10.1245.
КАС пабмед Статья Google Scholar

Bauer B, Hartz AM, Miller DS: Фактор некроза опухоли альфа и эндотелин-1 увеличивают экспрессию P-гликопротеина и транспортную активность через гематоэнцефалический барьер. Мол Фармакол. 2007, 71: 667-675. 10.1124/моль.106.029512.
КАС пабмед Статья Google Scholar
Hartz AM, Bauer B, Fricker G, Miller DS: быстрая модуляция опосредованного P-гликопротеином транспорта через гематоэнцефалический барьер с помощью фактора некроза опухоли-альфа и липополисахарида.Мол Фармакол. 2006, 69: 462-470. 10.1124/моль.105.017954.
КАС пабмед Статья Google Scholar
Hartz AM, Bauer B, Block ML, Hong JS, Miller DS: Частицы выхлопных газов дизельных двигателей вызывают окислительный стресс, провоспалительную сигнализацию и активацию P-гликопротеина на гематоэнцефалическом барьере. FASEB J. 2008, 22: 2723-2733. 10.1096/fj.08-106997.
Центральный пабмед КАС пабмед Статья Google Scholar

Block ML, Wu X, Pei Z, Li G, Wang T, Qin L, Wilson B, Yang J, Hong JS, Veronesi B: частицы дизельного выхлопа нанометрового размера избирательно токсичны для дофаминергических нейронов: роль микроглии, фагоцитоз, и НАДФН-оксидазы. FASEB J. 2006, 18: 1618-1620.
Google Scholar
Tai LM, Loughlin AJ, Male DK, Romero IA: P-гликопротеин и белок устойчивости к раку молочной железы ограничивают апикально-базолатеральную проницаемость эндотелия головного мозга человека для бета-амилоида.J Cereb Blood Flow Metab. 2009, 29: 1079-1083. 10.1038/jcbfm.2009.42.
КАС пабмед Статья Google Scholar
Xiong H, Callaghan D, Jones A, Bai J, Rasquinha I, Smith C, Pei K, Walker D, Lue LF, Stanimirovic D, Zhang W: ABCG2 активируется в мозге Альцгеймера с церебральной амилоидной ангиопатией и может действовать как привратник на гематоэнцефалическом барьере для пептидов Ab(1-40). Дж. Нейроски. 2009, 29: 5463-5475. 10.1523/JNEUROSCI.5103-08.2009.
Дж. Нейроски. 2009, 29: 5463-5475. 10.1523/JNEUROSCI.5103-08.2009.
Центральный пабмед КАС пабмед Статья Google Scholar
Vogelgesang S, Cascorbi I, Schroeder E, Pahnke J, Kroemer HK, Siegmund W, Kunert-Keil C, Walker LC, Warzok RW: Отложение бета-амилоида при болезни Альцгеймера обратно пропорционально экспрессии P-гликопротеина в мозг пожилых людей без деменции. Фармакогенетика. 2002, 12: 535-541. 10.1097/00008571-200210000-00005.
КАС пабмед Статья Google Scholar
Wijesuriya HC, Bullock JY, Faull RL, Hladky SB, Barrand MA: переносчики оттока ABC в сосудах головного мозга у пациентов с болезнью Альцгеймера. Мозг Res. 2010, 1358: 228-238. 10.1016/j.brainres.2010.08.034.
КАС пабмед Статья Google Scholar
Abbott NJ: Доказательства объемного потока интерстициальной жидкости головного мозга: значение для физиологии и патологии. Нейрохим Инт. 2004, 45: 545-552. 10.1016/j.neuint.2003.11.006.
Нейрохим Инт. 2004, 45: 545-552. 10.1016/j.neuint.2003.11.006.
КАС пабмед Статья Google Scholar
Cserr HF, DePasquale M, Patlak CS, Pullen RGL: Конвекция интерстициальной жидкости головного мозга и ее роль в регуляции объема мозга. Энн Н.Ю. Академия наук. 1989, 481: 123-134. 10.1111/j.1749-6632.1986.tb27144.x.
Артикул Google Scholar
Форбродт А.В.: Ультраструктурная цитохимия эндотелия гематоэнцефалического барьера. Прог Гистохим Цитохим. 1988, 18: 1-99.
КАС пабмед Статья Google Scholar
Zlokovic BV, Mackic JB, Wang L, McComb JG, McDonough A: Дифференциальная экспрессия изоформ альфа- и бета-субъединиц Na, K-АТФазы на гематоэнцефалическом барьере и сосудистом сплетении. Дж. Биол. Хим. 1993, 268: 8019-8025.
КАС пабмед Google Scholar

Lingrel JB: Физиологическое значение сайта связывания кардиотонического стероида/уабаина Na, K-АТФазы. Annu Rev Physiol. 2010, 72: 395-412. 10.1146/аннурев-физиол-021909-135725.
Центральный пабмед КАС пабмед Статья Google Scholar
O’Donnell ME, Lam TI, Tran LQ: Эстрадиол снижает активность котранспортера Na-K-Cl через гематоэнцефалический барьер и уменьшает образование отека при постоянной окклюзии средней мозговой артерии.J Cereb Blood Flow Metab. 2006, 26: 1234-1249.
ПабМед Статья КАС Google Scholar
Yang T, Roder KE, Bhat GJ, Thekkumkara TJ, Abbruscato TJ: Члены семейства протеинкиназ C в качестве мишени для регуляции гематоэнцефалического барьера Na, K, 2Cl-котранспортера во время инсульта in vitro и никотина воздействие. Фарм Рез. 2006, 23: 291-302. 10.1007/s11095-005-9143-2.
КАС пабмед Статья Google Scholar

Millar ID, Brown PD: NBCe2 демонстрирует стехиометрию 3 HCO3-:1 Na+ в эпителиальных клетках сосудистого сплетения мыши. Biochem Biophys Res Commun. 2008, 373: 550-554. 10.1016/j.bbrc.2008.06.053.
КАС пабмед Статья Google Scholar
Taylor CJ, Nicola PA, Wang S, Barrand MA, Hladky SB: транспортеры, участвующие в регуляции внутриклеточного pH в первично культивируемых эндотелиальных клетках головного мозга крысы. Дж. Физиол. 2006, 576: 769-785.10.1113/jphysiol.2006.117374.
Центральный пабмед КАС пабмед Статья Google Scholar
Vigne P, Farre AL, Frelin C: Na+-K+-Cl-котранспортер эндотелиальных клеток капилляров головного мозга. Дж. Биол. Хим. 1994, 269: 19925-19930.
КАС пабмед Google Scholar
Никола П.А., Тейлор С.Дж., Ван С., Барранд М.А., Хладки С. Б.: Транспортная активность, участвующая в восстановлении внутриклеточного рН после воздействия кислоты и щелочи в микрососудистых эндотелиальных клетках головного мозга крысы.Арка Пфлюгера. 2008, 456: 801-812. 10.1007/s00424-007-0441-х.
Б.: Транспортная активность, участвующая в восстановлении внутриклеточного рН после воздействия кислоты и щелочи в микрососудистых эндотелиальных клетках головного мозга крысы.Арка Пфлюгера. 2008, 456: 801-812. 10.1007/s00424-007-0441-х.
КАС пабмед Статья Google Scholar
Johanson CE, Duncan JA, Klinge PM, Brinker T, Stopa EG, Silverberg GD: Множественность функций спинномозговой жидкости: новые проблемы в области здоровья и болезней. Цереброспинальная жидкость Res. 2008, 5: 10-10.1186/1743-8454-5-10.
Центральный пабмед пабмед Статья КАС Google Scholar
Pollay M, Curl F: Секреция спинномозговой жидкости эпендимой желудочков кролика. Am J Physiol. 1967, 213: 1031-1038.
КАС пабмед Google Scholar
Damkier HH, Brown PD, Praetorius J: Эпителиальные пути транспорта электролитов сосудистого сплетения. Физиология (Bethesda). 2010, 25: 239-249.
Физиология (Bethesda). 2010, 25: 239-249.
КАС Статья Google Scholar
Praetorius J, Nejsum LN, Nielsen S: Продукт гена SCL4A10 избирательно картируется в базолатеральной плазматической мембране эпителиальных клеток сосудистого сплетения. Am J Physiol Cell Physiol. 2004, 286: C601-C610. 10.1152/ajpcell.00240.2003.
КАС пабмед Статья Google Scholar
Jacobs S, Ruusuvuori E, Sipila ST, Haapanen A, Damkier HH, Kurth I, Hentschke M, Schweizer M, Rudhard Y, Laatikainen LM, Tyynela J, Praetorius J, Voipio J, Hübner CA: Мыши с целевое нарушение гена Slc4a10 имеет маленькие желудочки головного мозга и снижает возбудимость нейронов.Proc Natl Acad Sci USA. 2007, 105: 311-316. 10.1073/пнас.0705487105.
Центральный пабмед пабмед Статья Google Scholar
Praetorius J, Nielsen S: Распределение переносчиков натрия и аквапорина-1 в сосудистом сплетении человека. Am J Physiol Cell Physiol. 2006, 291: C59-C67. 10.1152/ajpcell.00433.2005.
Am J Physiol Cell Physiol. 2006, 291: C59-C67. 10.1152/ajpcell.00433.2005.
КАС пабмед Статья Google Scholar
Damkier HH, Prasad V, Hubner CA, Praetorius J: Nhe1 представляет собой люминальный обменник Na+/H+ в сосудистом сплетении мышей и нацелен на базолатеральную мембрану у Ncbe/Nbcn2-нулевых мышей. Am J Physiol Cell Physiol. 2009, 296: C1291-C1300. 10.1152/ajpcell.00062.2009.
Центральный пабмед КАС пабмед Статья Google Scholar
Keep RF, Xiang J, Betz AL: Котранспорт калия в сосудистом сплетении крысы. Am J Physiol Cell Physiol.1994, 267: C1616-C1622.
КАС Google Scholar
Хьюз А.Л., Пахомова А., Браун П.Д.: Регуляторное увеличение объема эпителиальных клеток, выделенных из сосудистого сплетения четвертого желудочка мыши, включает обмен Na+-H+, но не Na+-K+-2Cl-котранспорт. Мозг Res. 2010, 1323: 1-10. 10.1016/j.brainres.2009.12.094.
Мозг Res. 2010, 1323: 1-10. 10.1016/j.brainres.2009.12.094.
КАС пабмед Статья Google Scholar
Redzic ZB, Isakovic AJ, Misirlic Dencic ST, Popadic D, Segal MB: Неравномерное распределение переносчиков нуклеозидов и внутриклеточная ферментативная деградация предотвращают транспорт интактного [ 14 C] аденозина через эпителий сосудистого сплетения овец в виде монослоя в первичной культуре. Цереброспинальная жидкость Res. 2006, 3: 4-10.1186/1743-8454-3-4.
Центральный пабмед пабмед Статья КАС Google Scholar
CDC – Малярия – О малярии
Комары Anopheles
Малярия передается человеку самками комаров рода Anopheles. Самки комаров потребляют пищу из крови для производства яиц, и эта кровь является связующим звеном между человеком и комаром-хозяином в жизненном цикле паразита. Успешное развитие малярийного паразита в комаре (от стадии «гаметоцитов» до стадии «спорозоитов») зависит от нескольких факторов. Наиболее важными являются температура и влажность окружающей среды (более высокие температуры ускоряют рост паразита в комаре), а также то, выживает ли Anopheles достаточно долго, чтобы позволить паразиту завершить свой цикл в комаре-хозяине («спорогонический» или «внешний» цикл). продолжительность от 9 до 18 дней).В отличие от хозяина-человека, хозяин-комар не страдает заметно от присутствия паразитов.
Наиболее важными являются температура и влажность окружающей среды (более высокие температуры ускоряют рост паразита в комаре), а также то, выживает ли Anopheles достаточно долго, чтобы позволить паразиту завершить свой цикл в комаре-хозяине («спорогонический» или «внешний» цикл). продолжительность от 9 до 18 дней).В отличие от хозяина-человека, хозяин-комар не страдает заметно от присутствия паразитов.
Схема взрослой самки комара
Карта мира, показывающая распространение преобладающих переносчиков малярии
Anopheles freeborni кровососущий комар
Увеличенное изображение
Общая информация
Существует около 3500 видов комаров, объединенных в 41 род. Малярия человека передается только самками рода Anopheles .Из примерно 430 видов Anopheles только 30-40 являются переносчиками малярии (т. е. являются «переносчиками») в природе. Остальные либо редко кусают людей, либо не выдерживают развития малярийных паразитов.
Географическое распространение
Anophelines встречаются по всему миру, кроме Антарктиды. Малярия передается различными видами Anopheles в разных географических регионах. В пределах географических регионов разные среды поддерживают разные виды.
Анофелины, которые могут передавать малярию, встречаются не только в эндемичных по малярии районах, но и в районах, где малярия ликвидирована.Таким образом, эти районы подвержены риску повторного заноса болезни.
Этапы жизни
Как и все комары, комары Anopheles проходят четыре стадии в своем жизненном цикле: яйцо, личинка, куколка и взрослая особь. Первые три стадии водные и длятся 7-14 дней, в зависимости от вида и температуры окружающей среды. Кусающая самка комара Anopheles может быть переносчиком малярии. Самцы комаров не кусаются, поэтому не могут передавать малярию или другие заболевания. Взрослые самки, как правило, живут недолго, и лишь небольшая их часть живет достаточно долго (более 10 дней в тропических регионах), чтобы передать малярию.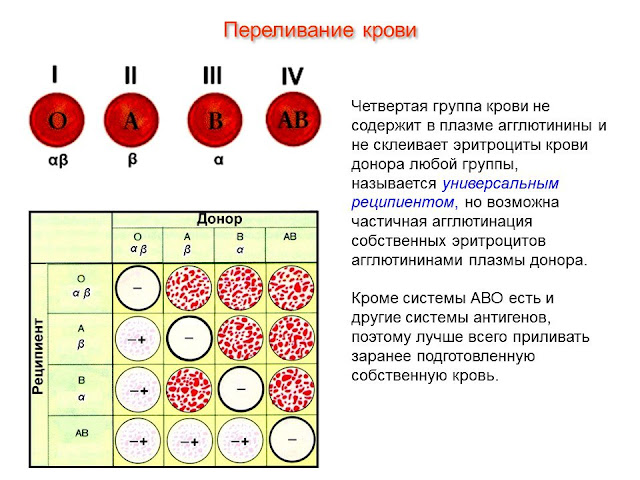
Яйца
Взрослые самки откладывают 50-200 яиц за одну кладку. Яйца откладываются поодиночке прямо на воду и уникальны тем, что имеют поплавки с обеих сторон. Яйца неустойчивы к высушиванию и вылупляются в течение 2-3 дней, хотя в более холодном климате вылупление может занять до 2-3 недель.
Личинки
Личинки комаров имеют хорошо развитую голову с ротовыми щеточками, используемыми для питания, крупную грудную клетку и сегментированное брюшко. У них нет ног. В отличие от других комаров, личинки Anopheles лишены дыхательного сифона и по этой причине располагаются так, что их тело находится параллельно поверхности воды.
Вверху: Анофелес Яйцо; обратите внимание на боковые поплавки.
Внизу: Anopheles яиц откладываются поодиночке.
Личинки дышат дыхальцами, расположенными на 8-м сегменте брюшка, поэтому должны часто всплывать на поверхность.
Личинки большую часть времени питаются водорослями, бактериями и другими микроорганизмами в поверхностном микрослое. Они делают это, поворачивая голову на 180 градусов и питаясь снизу микрослоя. Личинки ныряют под поверхность, только если их потревожить.Личинки плавают либо судорожными движениями всего тела, либо за счет движения ротовыми щетками.
Они делают это, поворачивая голову на 180 градусов и питаясь снизу микрослоя. Личинки ныряют под поверхность, только если их потревожить.Личинки плавают либо судорожными движениями всего тела, либо за счет движения ротовыми щетками.
Личинки проходят 4 стадии развития, или возрастные стадии, после чего они превращаются в куколок. В конце каждого возраста личинки линяют, сбрасывая свой экзоскелет или кожу, чтобы обеспечить дальнейший рост.
Anopheles Личинка. Обратите внимание на положение, параллельное поверхности воды.
Личинки встречаются в самых разных средах обитания, но большинство видов предпочитают чистую, незагрязненную воду.Личинки комаров Anopheles были обнаружены в болотах с пресной или соленой водой, мангровых зарослях, рисовых полях, травянистых канавах, берегах ручьев и рек, а также в небольших временных дождевых лужах. Многие виды предпочитают местообитания с растительностью. Другие предпочитают места обитания, в которых их нет. Некоторые размножаются в открытых, освещенных солнцем водоемах, в то время как другие встречаются только в затененных местах размножения в лесах. Несколько видов размножаются в дуплах деревьев или пазухах листьев некоторых растений.
Другие предпочитают места обитания, в которых их нет. Некоторые размножаются в открытых, освещенных солнцем водоемах, в то время как другие встречаются только в затененных местах размножения в лесах. Несколько видов размножаются в дуплах деревьев или пазухах листьев некоторых растений.
Куколка имеет форму запятой, если смотреть сбоку.Это переходная стадия между личинкой и взрослой особью. Куколки не питаются, но претерпевают коренные метаморфозы. Голова и грудь сливаются в головогрудь, а брюшко изгибается под ним. Как и в случае с личинками, куколки должны часто всплывать на поверхность, чтобы дышать, что они делают через пару дыхательных труб на головогруди. Через несколько дней после куколки дорсальная поверхность головогруди расщепляется, и взрослый комар выходит на поверхность воды.
Продолжительность от яйца до взрослой особи значительно различается у разных видов и сильно зависит от температуры окружающей среды.Комары могут развиваться от яйца до взрослой особи всего за 7 дней, но в тропических условиях обычно это занимает 10-14 дней.
Анофелес Куколка
Анофелес Взрослые. Обратите внимание (нижний ряд) на типичное положение покоя.
Взрослые
Как и все комары, взрослые анофелы имеют стройное тело с 3 отделами: головой, грудью и брюшком.
Голова предназначена для получения сенсорной информации и питания. На голове расположены глаза и пара длинных многочлениковых усиков.Усики важны для обнаружения запахов хозяина, а также запахов водных мест обитания личинок, где самки откладывают яйца. На голове также есть удлиненный, выступающий вперед хоботок, используемый для кормления, и два сенсорных щупика.
Грудная клетка предназначена для передвижения. К груди прикрепляются три пары ног и одна пара крыльев.
Брюшко приспособлено для переваривания пищи и развития яиц. Эта сегментированная часть тела значительно расширяется, когда самка принимает пищу крови.Кровь со временем переваривается и служит источником белка для производства яиц, которые постепенно заполняют брюшную полость.
Комаров Anopheles можно отличить от других комаров по длине щупиков, равной длине хоботка, и по наличию дискретных блоков черных и белых чешуек на крыльях. Взрослых особей Anopheles также можно определить по их типичному положению в состоянии покоя: самцы и самки отдыхают так, что их брюшки торчат вверх, а не параллельно поверхности, на которой они отдыхают .
Взрослые комары обычно спариваются в течение нескольких дней после выхода из стадии куколки. У некоторых видов самцы образуют большие стаи, обычно в сумерках, а самки прилетают в стаи для спаривания. Места спаривания многих видов остаются неизвестными.
Самцы живут около недели, питаясь нектаром и другими источниками сахара. Самки также питаются источниками сахара для получения энергии, но обычно им требуется кровяная мука для развития яиц. Получив полноценную кровяную муку, самка отдыхает несколько дней, пока переваривается кровь и развиваются яйца.Этот процесс зависит от температуры, но в тропических условиях обычно занимает 2-3 дня. Как только яйца полностью развились, самка откладывает их, а затем ищет кровь, чтобы выдержать еще одну партию яиц.
Как только яйца полностью развились, самка откладывает их, а затем ищет кровь, чтобы выдержать еще одну партию яиц.
Цикл повторяется до тех пор, пока самка не умрет. Самки могут прожить до месяца (или дольше в неволе), но большинство из них не живут дольше 1-2 недель в природе. Их шансы на выживание зависят от температуры и влажности, а также от их способности успешно питаться кровью, избегая защиты хозяина.
Самка Anopheles dirus кормление
Факторы, участвующие в передаче малярии и борьбе с малярией
Понимание биологии и поведения комаров Anopheles может помочь в разработке соответствующих стратегий борьбы. Факторы, влияющие на способность комара передавать малярию, включают его врожденную восприимчивость к Plasmodium , выбор хозяина и его продолжительность жизни. Наиболее опасны долгоживущие виды, предпочитающие человеческую кровь и поддерживающие развитие паразитов.Факторы, которые следует учитывать при разработке программы борьбы, включают восприимчивость малярийных комаров к инсектицидам и предпочтительное место кормления и отдыха взрослых комаров.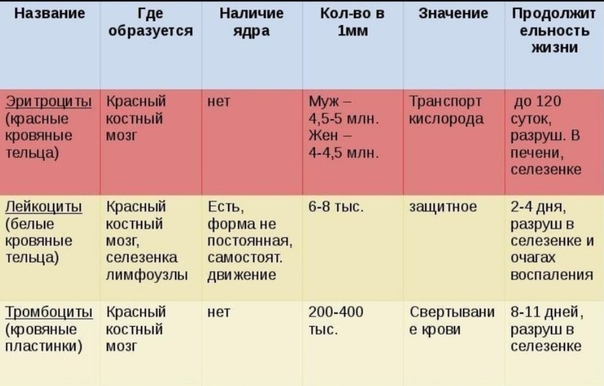
Подробнее о том, как уменьшить воздействие малярии
Предпочтительные источники крови
Одним из важных поведенческих факторов является степень, в которой виды Anopheles предпочитают питаться людьми (антропофилия) или животными, такими как крупный рогатый скот (зоофилия). Антрофильные Anopheles с большей вероятностью передают малярийных паразитов от одного человека к другому.Большинство комаров Anopheles не являются исключительно антропофильными или зоофильными; многие из них оппортунистичны и питаются любым доступным хозяином. Однако первичные переносчики малярии в Африке, An. gambiae и An. funestus , являются сильно антропофильными и, следовательно, являются двумя наиболее эффективными переносчиками малярии в мире.
Срок службы
После попадания в организм комара малярийные паразиты должны пройти развитие внутри комара, прежде чем они станут заразными для человека.Время, необходимое для развития комара (внешний инкубационный период), составляет 9 дней и более в зависимости от вида паразита и температуры. Если комар не выживает дольше, чем внешний инкубационный период, то он не сможет передавать каких-либо малярийных паразитов.
Если комар не выживает дольше, чем внешний инкубационный период, то он не сможет передавать каких-либо малярийных паразитов.
Непосредственно измерить продолжительность жизни комаров в природе невозможно, но многие исследования косвенно измеряли продолжительность жизни путем изучения их репродуктивного статуса или путем маркировки, выпуска и повторной поимки взрослых комаров.Большинство комаров не живут достаточно долго, чтобы передавать малярию, но некоторые могут жить в природе до трех недель. Хотя данные свидетельствуют о том, что уровень смертности увеличивается с возрастом, большинство работников оценивают продолжительность жизни с точки зрения вероятности того, что комар проживет один день. Обычно эти оценки варьируются от 0,7 до 0,9. Если выживаемость составляет 90% в день, то значительная часть населения будет жить дольше 2 недель и будет способна передавать малярию.Любая мера контроля, сокращающая среднюю продолжительность жизни популяции комаров, снизит потенциал передачи инфекции. Таким образом, инсектициды не обязательно сразу убивают комаров, но могут быть эффективными, ограничивая их продолжительность жизни.
Таким образом, инсектициды не обязательно сразу убивают комаров, но могут быть эффективными, ограничивая их продолжительность жизни.
Схемы кормления и отдыха
Большинство комаров Anopheles ведут сумеречный образ жизни (активны в сумерках или на рассвете) или ведут ночной образ жизни (активны ночью). Некоторые комары Anopheles питаются в помещении (эндофаги), в то время как другие питаются на открытом воздухе (экзофаги).После кормления кровью некоторые комары Anopheles предпочитают отдыхать в помещении (эндофильные), в то время как другие предпочитают отдыхать на открытом воздухе (экзофильные). Укусы ночных эндофагов комаров Anopheles можно заметно уменьшить за счет использования надкроватных сеток, обработанных инсектицидами (ITN), или за счет улучшения конструкции жилья для предотвращения проникновения комаров (например, оконных сеток). С эндофильными комарами легко бороться, распыляя внутри помещений инсектициды остаточного действия. Напротив, экзофагические/экзофильные переносчики лучше всего контролируются за счет сокращения источников (разрушения мест обитания личинок).
Напротив, экзофагические/экзофильные переносчики лучше всего контролируются за счет сокращения источников (разрушения мест обитания личинок).
Устойчивость к инсектицидам
Меры борьбы на основе инсектицидов (например, опрыскивание помещений инсектицидами, ОИС) являются основным способом уничтожения комаров, кусающих в помещении. Однако после длительного воздействия инсектицида в течение нескольких поколений у комаров, как и у других насекомых, может развиться резистентность, способность выживать при контакте с инсектицидом. Поскольку комары могут иметь много поколений в год, высокий уровень устойчивости может возникнуть очень быстро. Устойчивость комаров к некоторым инсектицидам была зарегистрирована в течение нескольких лет после того, как инсектициды были внедрены.Существует более 125 видов комаров с задокументированной устойчивостью к одному или нескольким инсектицидам. Развитие устойчивости к инсектицидам, используемым для остаточного опрыскивания помещений, было основным препятствием во время Глобальной кампании по ликвидации малярии. Разумное использование инсектицидов для борьбы с комарами может ограничить развитие и распространение резистентности, особенно путем чередования различных классов инсектицидов, используемых для борьбы. Мониторинг резистентности необходим для предупреждения программ борьбы о переходе на более эффективные инсектициды.
Разумное использование инсектицидов для борьбы с комарами может ограничить развитие и распространение резистентности, особенно путем чередования различных классов инсектицидов, используемых для борьбы. Мониторинг резистентности необходим для предупреждения программ борьбы о переходе на более эффективные инсектициды.
Восприимчивость/Рефрактерность
Некоторые виды Anopheles являются плохими переносчиками малярии, поскольку паразиты плохо развиваются (или вообще не развиваются) внутри них. Существуют также различия внутри видов. В лаборатории удалось отобрать штаммы An. gambiae , устойчивые к заражению малярийными паразитами. Эти устойчивые штаммы имеют иммунный ответ, который инкапсулирует и убивает паразитов после того, как они вторглись в стенку желудка комара.Ученые изучают генетический механизм этой реакции. Есть надежда, что когда-нибудь генетически модифицированные комары, невосприимчивые к малярии, смогут заменить диких комаров, тем самым ограничив или ликвидировав передачу малярии.